File: <ascomycetes.htm> <Index to Mycology> Pooled References <Glossary> Site
Description <Navigate to
Home>
Page 1
True Fungi (Eumycophyta1
Ascomycota (Ascomycetes, Ascomycotina)
-- Sac Fungi
(Contact) Please CLICK on underlined
links & included illustrations for details Use Ctrl/F to search for Subject Matter:
Tables Plates
Sample Examinations CLICK on
illustrations to enlarge: Introduction All members of the Ascomycota
produce an ascus
that contains ascospores. The class includes the largest group and
most successful of all fungi, with over 44,000 known species. The group has existed for many millions of
years and there is some evidence that they took their origin from
Zygomycotous forms. The Ascomycota
are highly important in the break down of organic matter in the soil, as
plant pathogens and for the production of antibiotics and other industrial
substances. Many species are purely
saprophytic; some are obligate parasites and others facultatively saprophytic
or parasitic. Most species have a
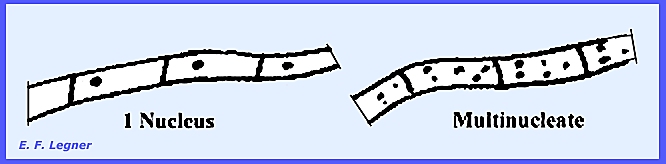
well-developed septate mycelium, and septations contain a septal pore or perforate septum. Cells primarily have only a single nucleus
per cell but there are some with more than one nucleus per cell.
Those species known as yeasts are
distinguished by not forming mycelia and individual cells multiply by fission
or budding processes. The yeasts are
not primitive organisms but special forms that evolved from a retrogressive
process in evolution. A large proportion of the
Ascomycetes have one or more means of vegetative reproduction. This imperfect stage produces conidia but
never any zygospores. The perfect or
sexual stage produces asci, which is the cardinal feature of the entire
class. The production of asci
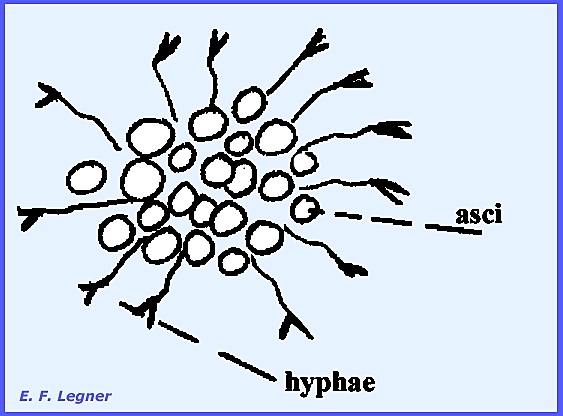
terminates the relatively complicated sexual process. Asci occur in a spore sac, which may take
various shapes.
The number of ascospores is
variable, but 8 per ascus are typical in 98 percent of species. Typically, ascospores are forcibly
discharged (sporangiospores are not).
The ascus is the seat of meiosis, which is not true in sporangia. In 99 percent of cases the ascus is also
the site of nuclear fusion. In contrast to sporangiospore
formation by progressive cleavage, ascospores are delimited in a process
known as "Free-cell Formation".
This results in a leaving of cytoplasm in the ascus, which is called epiplasm. Ascospores are almost always uninucleate
(haploid with a single nucleus per spore), but the genus Neurospora is
an exception with 2-4 nuclei per spore.
The epiplasm largely disappears when the ascospores mature, which
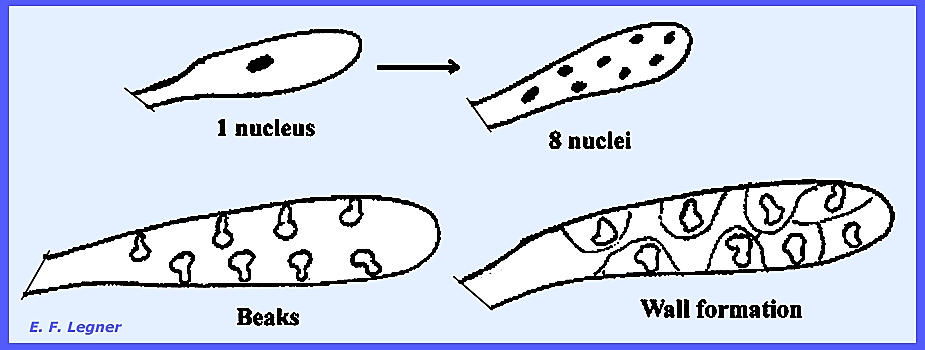
suggests that it serves to nurture the spores. As the ascus
develops there is a large 2N nucleus that divides to form 8 nuclei. These nuclei develop beaks. Cytoplasmic radiations form walls around
each nucleus thereby delimiting ascospores.
Fruiting
bodies (ascocarps)
whose walls consist of closely interwoven hyphae may form various
shapes. In contract there are no
fruiting bodies in the Zygomycota.
But not all
of the Ascomycota produce fruiting bodies.
There are none in the small sub-class Hemiascomycetes while all
members of the much larger subclass Euascomycetes produce fruiting bodies (=
ascocarps). ---------------------------------- Please
refer to the following plates for characteristic structures in the Ascomycota: Ascomycota Plate
102 = Conidiophore types: Phyllosticta,
Dendrophoma, Monopodium, Verticillium, Aspergillus, Penicillium & Isariopsis. Plate
103 = Asexual fruiting bodies: Septoria,
Marssonia, Epicoccum & Arthrobotryum. Plate
104 = Sexual reproduction & ascus
development in Ascomycota: Pyronema omphalodes. Plate
105 = Types of asci: Globose, Ovate, Septate, Clavate,
Cylindrical. Plate
106 = Variety of ascospores (20 types). Plate
107 = Four ways that Ascomycota bear asci. Plate
108 = A section thru' the stroma revealing
embedded ascocarps. Plate
109 = Several types of openings (pores) in
asci. Plate
110 = Four stages in ascospore
germination: Gelasinospora autosteira ---------------------------------- Two discussed orders in the Hemiascomycetes
are: Endomycetales
& Taphrinales
Three families in the order Endomycetales are: Ascoidaceae,
Endomycetaceae &
Spermophtoraceae. The Ascoidaceae
is well represented by
the Genus Dipodascus. Here the mycelium consists of
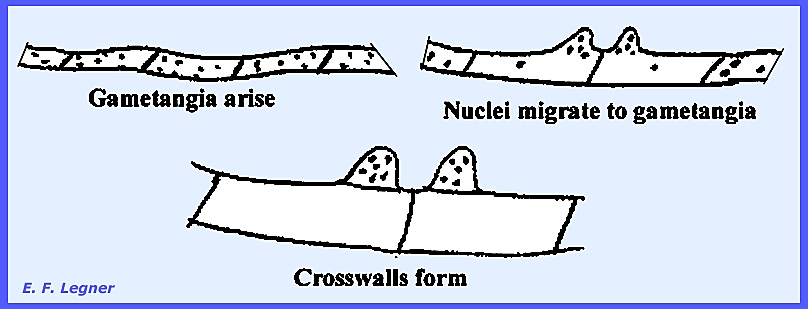
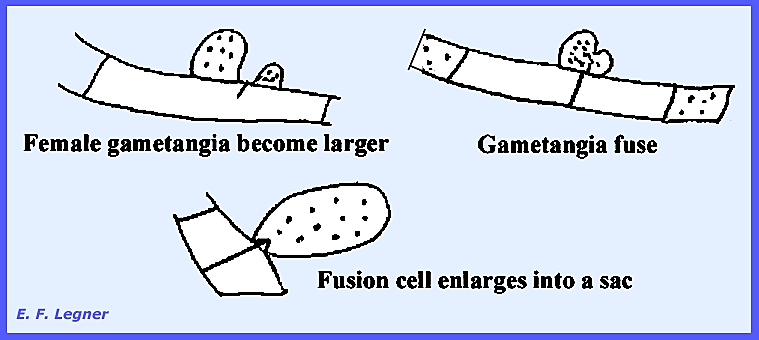
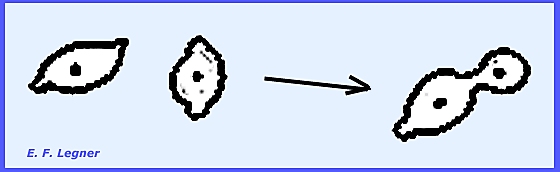
multinucleate cells and thee is no imperfect stage. Asci are formed as follows: Two gametangia arise on adjacent
cells and nuclei migrate to gametangia where crosswalls are formed.
The female portion usually
enlarges and gametangia fuse. The
fusion cell enlarges into a sac, which is a transformed fused gametangium
with many nuclei.
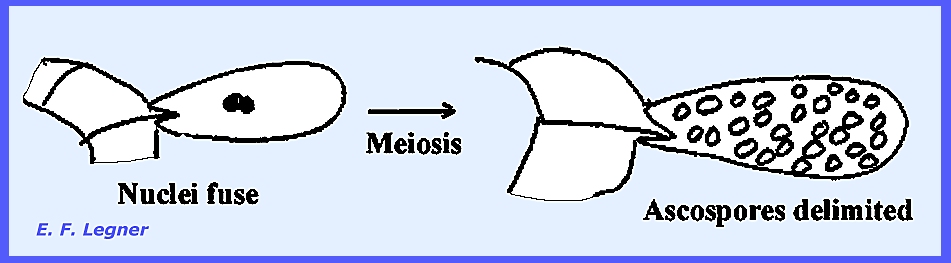
Fusion nuclei fuse but others
disintegrate. Meiosis takes place to
produce many nuclei. Ascospores are
delimited from each of the many nuclei.
----------------------- The family Endomycetaceae is
characterized by a septate mycelium and the absence of a fruiting body, all
the asci being scattered on the mycelium.
All species form asci as a result of direct fusion of gametangia; but
there is only a single nucleus per gametangium. Gametangia fuse and the nuclei fuse to produce one diploid
nucleus. Meiosis occurs and the
number of spores varies between 4 and 8.
There is no degeneration of the nuclei and the duration of the 2N
nucleus is short except in Saccharomyces cerevisiae. Various species show distinct
behaviors. In the Genus Eremascus
there is only a perfect stage.
But Endomyces
has an imperfect stage also. In
this species arthrospores are formed by disarticulation of hyphal cells. The process begins at the apex of hypha
that have stopped growing and continues posteriorly. Arthrospores germinate directly or they
may form crosswalls and multiply by fission.
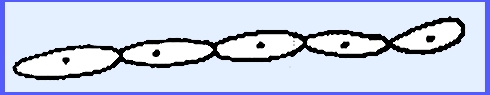
In Endomycopsis blastospores are formed in a budding
process. The blastospore gives rise
to hypha that may produce secondary blastospores and are thus difficult to
distinguish from yeasts.
Schizosaccharomyces and Saccharomyces are
yeasts that grow primarily in single-celled form. They multiply by fission.
In Schizosaccharomyces asci formation occurs when two cells come together
(gametangia), fusion occurs and eight ascospores are delimited in one ascus.
Saccharomyces is a budding
yeast, and S. cerevisiae is one of the most important species that is
used for rising bread. There are
individual cells and multiplication is accomplished by the formation of
buds. A single nucleus divides with
one nucleus going to the bud. In the
sexual stage every vegetative cell under the right conditions can form
ascospores within, and typically four are delimited. This process appears to be parthenogenetic
but it is not.
----------------------- Details on Characteristics of Yeasts Yeasts are not regarded as
primitive and they multiply either by fission or budding. The many different types are distinguished
as being sporogenous (produce asci), asporagenous (do not produce asci as in
the Deuteromycota), haplobiontic, budding, apiculate, bipolar, film-forming,
oxidative, diplobiontic, illegitimate diploid and multipolar. A typical haplobiontic yeast is Schizosaccharomyces
octosporus. A septum
separates the cell, and a cell may function as a gametangium, which fuse to
form an ascus. Fusion of nuclei
occurs in the developing ascus, and meiosis follows to produce 8 haploid
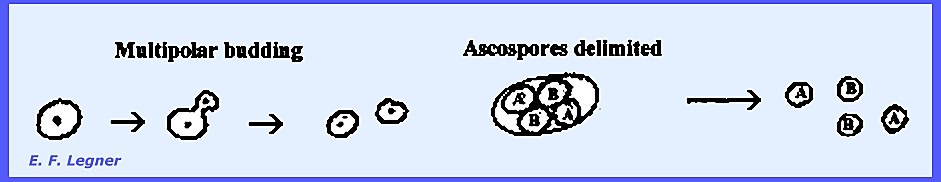
nuclei. An example of a budding (multipolar) yeast is Saccharomyces
cerevisiae. It is intermediate between haplobiontic and
diplobiontic and budding is the process of multiplication. It is a multipolar budding type where a
bud may appear on any side of the cell.
Diploid cells result from the budding process after fusion of
"A" and "B" asci.
Four ascospores are delimited in a cell (ascus).
Ascospores multiply by a budding
process, but are haploid and much smaller than the diploid cells. Under ordinary circumstances two of the
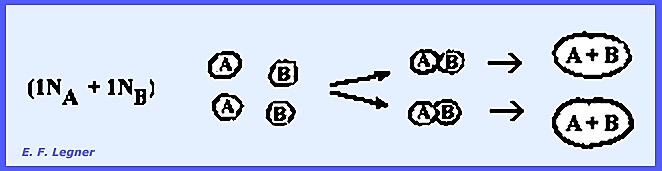
haploid cells come together and fuse.
Half of the ascospores in an ascus are of the "A" Type and
half of the "B" Type. When
two or one types fuse the result is an "illegitimate diploid." The budding of fused ascospores produces
diploid cells:
S. cerevisiae is of great
importance because under conditions of low aeration it can produce alcohol
without the intervention of Oxygen, even though the yeast cells will not
grow. C6H12O6 -----------► CO2 + C2H5OH Various strains are used in the
fermentation of beer, wine, etc. The
most tolerant strains will not tolerate more than 13-15 percent alcohol. Taxes from the sale of alcohol can surpass
6 billion dollars per year in North America, and the baking industry uses
yeast for leavening and carbon dioxide.
An estimated 300 tons of yeast cake is used daily in North America If Oxygen is present, the reaction
will be: C6H12O6
O2 -----------► CO2 + H2O This
reaction is used in the Baking and Yeast Production Industries. The medium is acidified and aerated to
stimulate rapid multiplication. In
about 12 hours the yeast will multiply to five times its original volume. Yeasts cannot use starch so they
must have sugar. Sugar sources are
usually molasses or grain mash (acid hydrolysis or enzyme or another
microorganism breaks down the starch into sugar). Yeasts are rich in protein and
there is a possibility of using sawdust, etc. as a food source for yeasts,
which are substituted for high protein diets. They are extensively used for livestock. Yeasts also are high in Vitamin B
complexes and Vitamin D (Ergosterol). Apiculate yeasts are exemplified
by the Genus Hanseniaspora,
which is found on ripening fruits (apple, grapes). They may also occur on dust, leaves in the soil, etc. These yeasts multiply in the juice and
carry on a kind of fermentation that is known as "Spontaneous
Generation Fermentation."
However, the alcohol content produced is low, in the range of 5-6
percent. In the commercial production
of wine, the juice is first sterilized and then inoculated with a desired
strain of yeast. The insect Genus Drosophila
feeds mainly on the yeasts although the initial attraction may involve the
alcohol. The apiculate yeasts bud
only at the poles and are thus "bipolar."
The Genus Pichia typifies film-forming yeasts. They have elongated cells in a chain known
as a pseudomycelium. They occur on
the surface of liquids such as pickle brine and are poor in the ability to
carry on alcoholic fermentation. In
some cases they may actually be exclusively oxidative.
Characteristics of the family Spermophthoraceae are
shown in the genera Spermophthora, Eremothecium and Ashbya. There is a mycelium and some species are
the cause of "Cotton Stigmatomycosis," which damages the cotton boll. The Genus Nematospora is
single-celled and causes "Yeast Spot of Pea."
All these genera produce Rhiboflavin in large amounts. Eremothecium exceeds all others in production, but
following the placing of a patent, several strains of Ashbya have been
found that almost equal it in production. ----------------------- Please refer to the
following plates for characteristic structures and Life Cycles in the Endomycetales: Ascomycota: Hemiascomycetes: Endomycetales Plate
50 = Ascomycota: Hemiascomycetes, Ascoidaceae: Dipodascus sp. Plate
111 = Life Cycle -- Dipodascus uninucleatus. Plate
112 = Life Cycle -- Eremascus fertilis. Plate
113 = A yeast cell showing various
structures. Plate
114 = Chain of yeast cells (pseudomycelium)
produced by budding. Plate
115 = Five types of yeast ascospores. Plate
116 = Life Cycles -- Schizosaccharomyces octosporus, Saccharomycodes ludwigii, Saccharomyces
cerevisiae Plate 185 = Life
Cycle -- Endomycetaceae: Endomyces sp. & Endomycopsis sp. Plate 186 = Life
Cycle -- Endomycetaceae: Schizosaccharomyces octosporus Plate 187 = Life
Cycle -- Endomycetaceae: Saccharomyces cerevisiae; &
Structures of Hanseniaspora sp & Pichia sp. Plate 189 = Example
Structures: Endomycetales: Endomyces
sp., Schizosaccharomyces
octosporus & Endomycopsis sp. Plate 190 = Example
Structures: Endomycetales: Saccharomyces
sp. & Ashbya gossypii. ----------------------- The order Taphrinales
is represented here by two families: Taphrinaceae and Protomycetaceae. Asci arise from a proascus, not from two
gametangia as in the Endomycetales.
No fruiting bodies are present. The family Taphrinaceae is represented by a
single genus, Taphrina, which causes various plant diseases. Taphrina deformans causes
"Peach
Leaf Curl", T. communis and T. pruni cause
"Plum
Pockets." Also
various species of Taphrina occur on many wild hosts of which about
one-third are pathogens on ferns. A naked layer, hymenium, bears asci on the
surface. They may be subcuticular,
breaking the host cuticle on emergence or they may reside in the epidermal
cell interspaces. Each arises from a proascus. The mycelium is intercellular (= "High Type"
parasite) and there are no haustoria.
All cells have two nuclei (= dikaryotic mycelium), and this
is the only place in all the Ascomycota where this occurs. In the
Basidiomycota a dikaryotic mycelium is common; thus, the similarity. Ascospore budding may begin inside the
ascus, which resembles a yeast on agar (Saccharomyces).
Taphrina deformans exemplifies
the family. Hyphae accumulate
underneath the host cuticle and are called proasci.
Nuclei fuse in the proascus to
give a diploid condition.
The diploid cell pushes up through
the host cuticle and forms a crosswall.
The bottom cell is now the "foot cell" and the top one
the ascus.
Meiosis occurs in the ascus and
subsequent mitotic divisions produce an ascus with 8 haploid nuclei.
Each nucleus forms an ascospore
and these may be forcibly discharged from the ascus.
The ascospores bud until reaching
an optimum substrate, where they stop budding and send out a germ tube
between the epidermal cells. Hyphae
may occur throughout the leaf tissue.
Taphrina deformans is a
homothallic organism and there is no imperfect stage. There are at least two heterothallic
species of Taphrina in Europe.
In this case "A" and "B" ascospores are
produced. As "A" and "B" ascospores fuse and
"A & B" nucleus is formed. Other differences among the
species of Taphrina may include the presence of foot cells, and the
size of the ascus varies (e.g., T. coerulescens are very large). Budding may also occur in the ascus. Taphrina species may affect
the host by causing hypertrophy with or without hyperplasia (e.g., ferns show
little hypertrophy). There may be a
failure of tissue differentiation as in the case of the disease known as
"Plum Pockets." ---------------------------------- The family Protomycetaceae is typified by the
Genus Protomyces. These are plant invaders that incite gall
production on ragweed. Thick-walled
cells arise on the mycelium (= proasci).
Intervening hyphae break down
leaving only the proasci. These are
located down in the plant tissue.
Ascospores bud on being released from the ascus. However, there is some doubt as to whether
or not these are proasci. Cytological
behavior in the "ascus" is quite unlike that of other members of
the Ascomycota.
----------------------- Please refer to the following plates for characteristic structures
and Life Cycles in the Taphrinales: Ascomycota:
Hemiascomycetes: Taphrinales Plate
117 = Life Cycle-1 -- Taphrina deformans. Plate 188 = Life
Cycle-2 -- Taphrinaceae: Taphrina deformans "Peach Leaf Curl." Plate 191 = Plant
Host Symptoms -- Taphrinales: Taphrina spp. Plate 192 = Example
Structures: Ascomycota: Taphrinales ----------------------------- In the Sub-Class Euascomycetes,
Series: Plectomycetes
three orders discussed are:
Plectascales, Myriangiales and Erysiphales. In the Series: Pyrenomycetes seven orders discussed are
Hypocreales, Sphaeriales, Pseudosphaeriales, Dothideales, Hemisphaeriales,
Laboulbeniales and Hysteriales. In
the Series: Discomycetes four orders are Helotiales, Lecanorales, Pezizales
and Tuberales. The Euascomycetes show different
developmental patterns, but are generally similar throughout. Asci are produced in aggregates and formed
in connection with a fruiting body, the ascocarp. A sexual process involving a female ascogonium and a male antheridium
or spermatium initiates asci formation.
Gametangia fuse followed by a transfer of nuclear material to the
ascogonium. The ascogonium gives rise
to ascogenous hyphae in which nuclei are paired (= dikaryon). Conjugate nuclear division is carried out
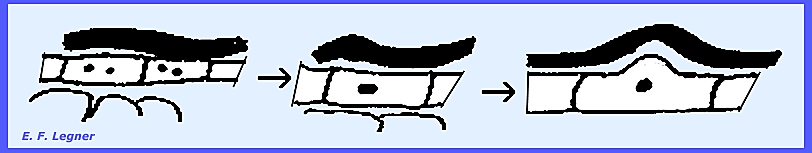
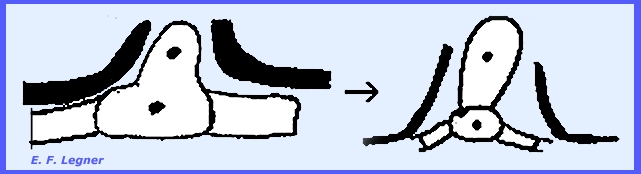
in the ascogenous hyphae and these hyphae re dikaryotic. A ascogenous hypha branch bends over in a
crozier. Three kinds of cells are
formed: (1) ultimate, (2) penultimate
and (3) antepenultimate. The penultimate cell becomes the ascus.
The ultimate cell may fuse with
the antepenultimate cell to form a dikaryon and another crozier. This may then form another ascus.
Sterile hyphae are induced to grow
and branch underneath the sexual structure.
These sterile structures either form the main part of the ascocarp or the
system of ascogenous hyphae may simply give rise to a structure of sterile
hyphae below the sexual structures, which is still an ascocarp, however.
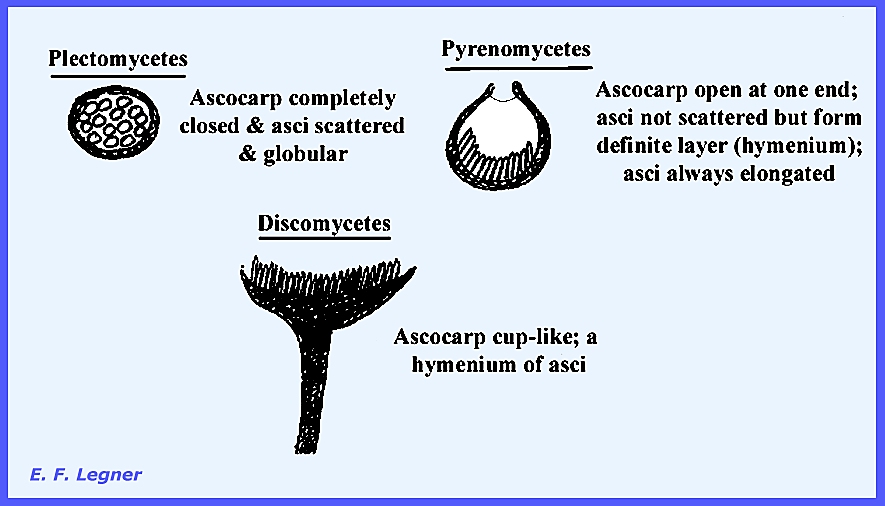
Three types of ascocarps are found
in the three respective groups of the Euascomycetes, or series (Fig 277).
------------------------------------ The order Plectascales is
characterized by the families Gymnoascaceae, Aspergillaceae and
Elaphomycetaceae In the Family Gymnoascaceae,
the Genus Byssochlamys
includes what are known as "Barnyard fungi." These are karotinophilic fungi that grow
on feathers, hair, hoofs, nails, maize, etc.
They are similar to Dermophytes in the Deuteromycota, and they can
produce a mild skin disease in humans that is not as serious as in the
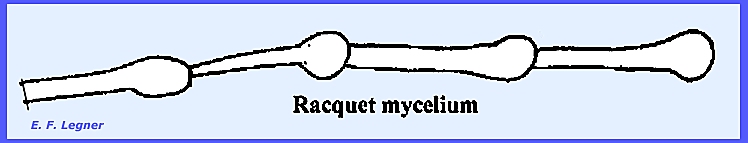
Deuteromyccota. They have a Racquet mycelium
where the hyphae are racquet-shaped.
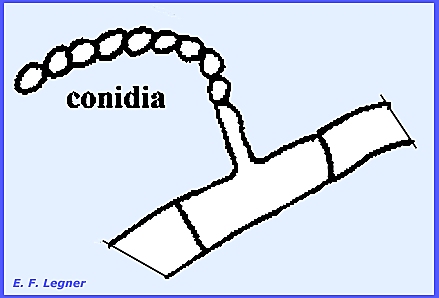
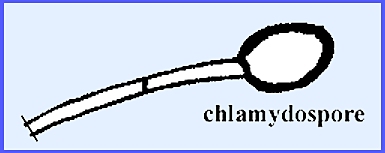
Two kinds of vegetative
reproduction are by conidia or chlamydospores. The conidia occur in long chains at the ends of branched or
unbranched conidiophores. The
Deuteromycota have a genus Paescilomyces that produces conidia in the
same manner.
During sexual reproduction
antheridia and ascogonia fuse.
Ascogenous hyphae branch out from the ascogonium and the ascus forms
as a result of crozier formation from the ascogenous hyphae. Many asci are produced, which are
typically naked, each with 8 ascospores.
There is no real ascocarp although there may be a weak suggestion of
one.
In the Genus Gymnoascus a sexual process
initiates the building the building up of a weak wall, and spines occur on
the tips of swelled hyphae. These are
considered to be a "weft of specialized hyphae."
-------------------------------------- Principal
Characteristics of Euascomycetes Asci are clustered and borne in a
fructification (ascocarp). The asci
are formed in connection with an ascogenous hyphal system. Ascogenous hyphae arise as outgrowths of
an ascogonium. Nuclei pass from an
antheridium to an ascogonium but do not fuse. Instead, conjugate nuclear division increases each nucleus
respective to set up a dikaryotic phase in the ascogenous hyphal system. The dikaryophase is ended when the asci
form, which involves a crozier. Classification of the groups
(series) is based on the form of the ascocarp, the form of the asci and the
distribution of asci in the ascocarp.
The Plectomycetes have a closed fructification (cleistothecium),
scattered asci and globular asci. The
Pyrenomycetes have a flask-shaped fructification (perithecium), asci occur in a
definite layer (hymenium) and they are elongated. The Discomycetes have an open fructification (apothecium),
asci occur in a definite layer and they are also elongated. The order Plectascales is
the most characteristic order in the group.
Most ascocarps are very small although the family Elaphomycetaceae is
an exception (some doubt about its classification). The family Gymnoascaceae
has poorly developed ascocarps (thin wefts of hyphae), but all other typical
features of Euascomycetes are present.
A scattered group of more or less globular asci each containing from
6-8 spores is formed. The genus Byssochlamys
has relatively undeveloped ascocarps.
The family also has karytinophilis forms. It is related to a group of Fungi Imperfecti (Dermatophytes)
ecause of the peculiarity of the racquet mycelium. ----------------------------------- The family Aspergillaceae has very small
ascocarps with well-defined peridial walls and a well-developed
cleistothecium. The asci are
globular, scattered in the ascocarp with 8 ascospores per ascus, and thus are
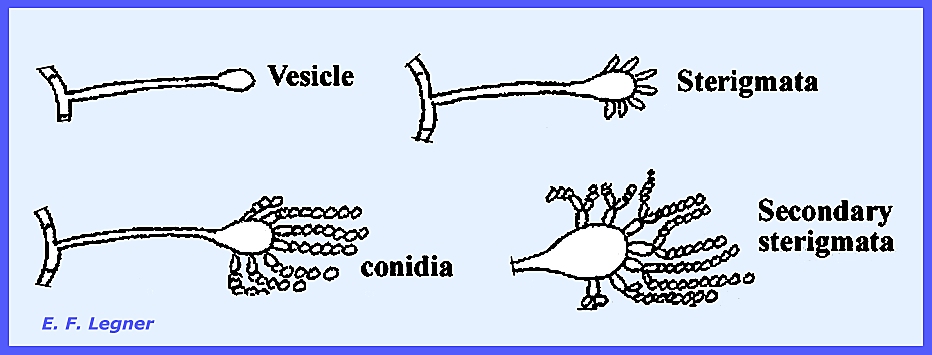
similar to the Gymnoascaceae. The Genus Aspergillus has a characteristic pattern in
the conidial stage. A conidiophore
arises anywhere on the mycelium and forms a bulbous structure at its apex (=
vesicle). Sterigmata are formed, and
conidia are produced in chains at the ends of the sterigmata. Sometimes there are secondary sterigmata
also. Similar forms in the Fungi
Imperfecti bear the name Aspergillus, but they never possess the added
names of Eurotium, Emericella, and Sartorya.
Members of the genus possess the
ability to grow where there is a high osmotic concentration (high sugar,
salt, etc.). Species that best
represent the genus are Aspergillus ametelodami, with both sexual and
asexual stages; and Aspergillus niger, A. fumigatus and A.
flavus that do not have a perfect stage. Examples of species with economic
importance are Aspergillus alliaceus (Bulb Rot of Onion), A. flavus
(starch-digesting enzyme), A. fumigatus (causes Aspergillosis), A.
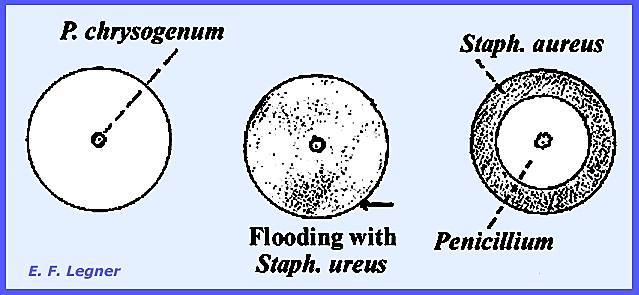
niger (produces citric acid.) The Genus Penicillium is a companion
genus of Aspergillus, both occurring in the soil. However, Penicillium exceeds Aspergillus
in abundance. The ascocarpic stage is
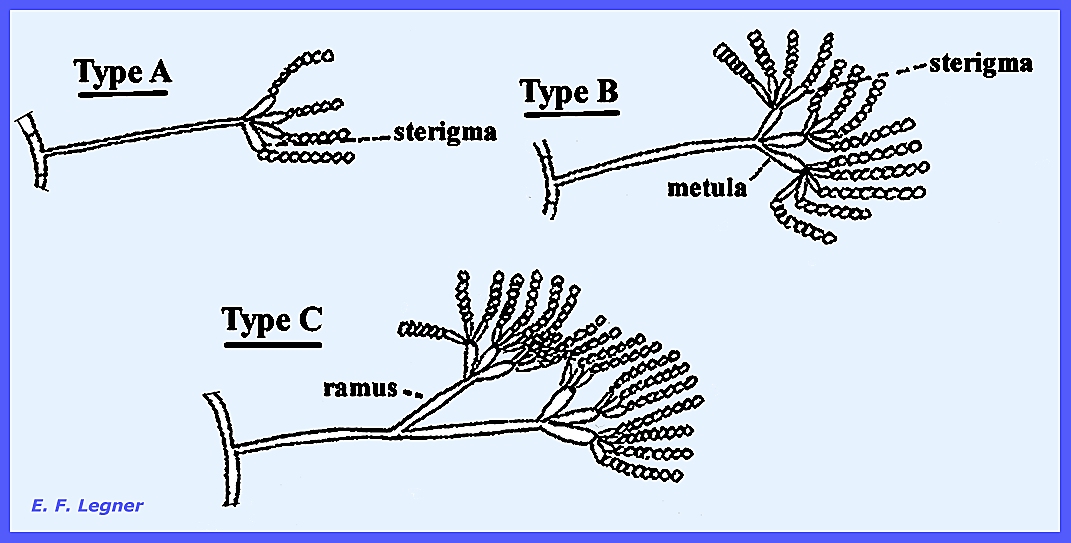
similar to Aspergillus, but the conidial stage differs. Here conidiophores do not form a vesicle,
instead a penicillus
of where there are three types (Fig 284).
The structure bearing the chains of conidia are called sterigmata.
The abundance of Penicillium
spp. In nature is extensive. They are
primarily saprophytic forms and plan an important role in the breakdown of
organic matter. They also may spoil
many foodstuffs. There is no perfect
stage. Penicillium chrysogenum
secretes penicillin. It acts on
gram-positive organisms, which are very common in nature. Although other species also produce
antibiotics, P. chrysogenum is widely used for antibiotics production
due to its secretions being low mammalian toxicity. Other species are P. expansum (Apple Rot), P.
digitatum and P. italicum (Citrus Fruit Rot), P. roqueforti
(Blue Cheeses). Thielavia basicola occurs in
the soil and was once erroneously thought to be pathogenic on tobacco. The disease "Thielaviopsis" was
actually caused by one of the Fungi Imperfecti. Ascocarps are similar to Aspergillus and Penicillium,
but they are dark and football shaped.
The ascospores spill out into the cavity of the cleistothecium and the
ascus is evanescent. Monascus spp. Cause the
disease "Purple silage mold."
This name was derived from a misjudgment, as it was once believed that
the scattered ascospores occurred in one large ascus. The family Elaphomycetaceae has
ascocarps far different from any previous form. They are the size of a hickory nut and hypogenous. They are associated with tree roots and
believed to be a mycorrhizal fungus by occurring on tree roots. The Genus Elaphomyces may actually be a highly
modified Discomycete.
----------------------------- Please refer to the
following plates for characteristic structures and Life Cycles in the Plectascales: Ascomycota: Euascomycetes: Plectascales Plate 118 = Conidiophores & hypha of
Aspergillus. Plate
119 = Cleistothecia cross sections: Aspergillus
sp. Plate
120 = Conidiophores: Penicillium
thomii, P. lanoso-coeruleum
& P. wortmanni. Plate
121 = Life Cycle -- Penicillium vermiculatum. Plate 193 = Life
Cycle -- Plectascales: Gymnoascaceae:
Byssochlamys sp. Plate 194 = Life Cycle
-- Plectascales: Aspergillaceae: Penicillium
vermiculatum Plate 195 = Life
Cycle -- Plectascales:
Aspergillaceae: Aspergillus sp. ----------------------------- The order Myriangiales has
short and wide asci, which are scattered.
There is no typical cleistothecium.
Species in the genus Myriangium are
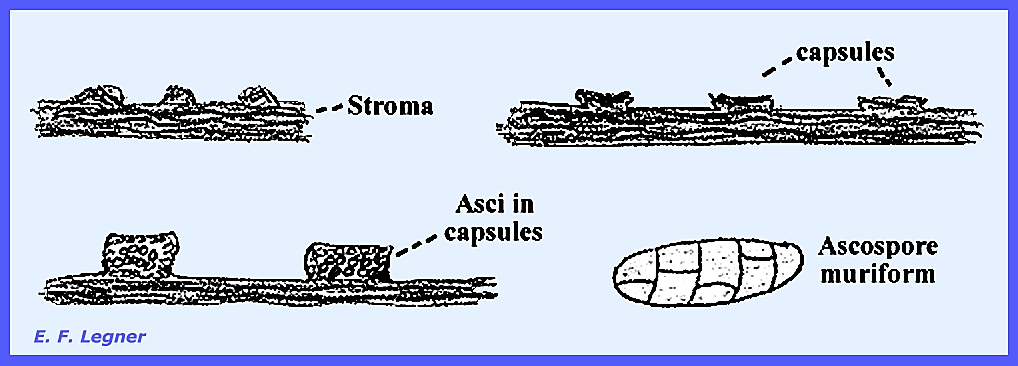
parasitic on scale insects. They
build a dense mass of compacted hyphae (stroma), which flatten out to form
capsules. The whole structure is stromatic. The asci are borne scattered in the
capsules. The ascospores are muriform
where multicellular spores appear as bricks in a wall.
The Genus Elsinoe also has a stromatic
form. Ascospores are scattered
through an undifferentiated stromatic layer.
The genus includes pathogens of cultivated plants and causes "Spot Anthracnose"
especially in tropical regions. The
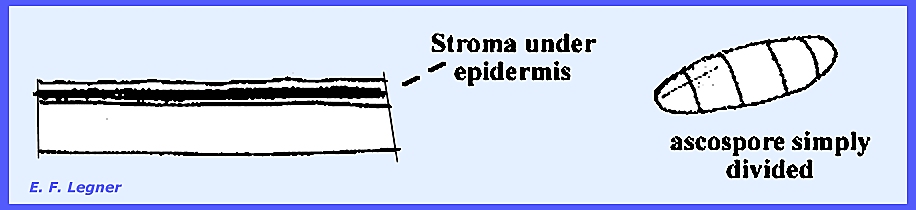
imperfect stages is referred to as "Sphaceloma. A stroma appears as a layer on or under
the host epidermis. Ascospores are
not muriform but are simply divided by a few crosswalls.
----------------------------- Please refer to the
following plates for characteristic structures and Life Cycles in the Myriangiales: Ascomycota:
Euascomycetes: Myriangiales Plate
122 = Life Cycle -- Elsinoe veneta. Plate
123 = Structures of Myriangium bambusae. Plate 196 = Life
Cycle -- Myriangiales: Elsinoe sp. ----------------------------- Please refer to the
following plates for characteristic structures and Life Cycles in the Plectascales
& Myriangiales: Plate 197 =
Plectascales & Myriangiales Example Structures: Aspergillus amstelodami,
Penicillium Byssochlamys nivea, carpenteles,
P. frequentans, Thielavia basicola. Plate 198 = Plectascales
& Myriangiales Example Structures: Aspergillus
fumigatus, Byssochlamys
nivea, Elaphomyces sp., Elsinoe wisconsinensis, Monascus sp.,
Myriangium sp. ----------------------------- The order Erysiphales, which
is almost equivalent to the group formerly known as
"Perisporiales", contains several families, the members of which
are usually found growing on leaves, stems and fruits of seed plants. The mycelium is largely, if not entirely,
confined to the surface of the suscept organ involved and may be either white
or dark colored. There are both
obligate parasites, as in the Erysiphaceae, or forms merely growing as
saprophytes on honeydew that is deposited by insects. The ascocarps are mostly cleistothecia. The family Erysiphaceae
includes the Powdery Mildews. All are common parasites. The mycelium is primarily superficial on
leaves, stems and even fruit, but may be internal also. Haustoria anchor the mycelium to
epithelial cells, which forms a whitish powdery mass with profusion of
conidia. The mycelium is composed of
short, uninucleate cells, and the haustoria are bulbous usually with a single
nucleus. They may be elongated or
they may even occur uncommonly in the subepidermal layer. The Meliolaceae and
Capnodiaceae have a dark superficial mycelium, and these forms are common in
the tropics.
The Erysiphaceae incite a great
variety of diseases of cultivated and wild plants. The grape industry in France was threatened before with the
surge of the Downy Mildews (Zygomycota).
Dusting with sulfur onto the vines successfully controlled the
infection that was the first successful use of a fungicide. But in humid areas the powdery mildews may
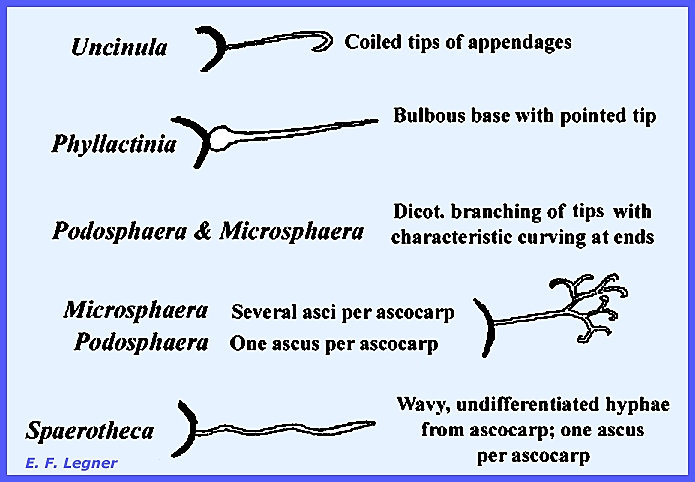
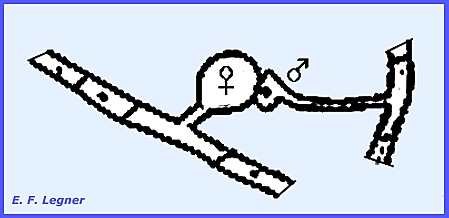
be reduced by moisture on the leaf surfaces. The Genus Phyllactinia is only one of
the six common genera in North America.
It has both a superficial and an internal mycelium. There is no form with only an internal
mycelium. There is an Oidium Imperfect
Stage, where the fungus bears unbranched, upright conidiophores
with catenulate conidia.
The conidiophore typically has
only one nucleus. The nucleus keeps
dividing in the conidiophore and migrates to the terminus to form a new
conidium with an indefinite number of conidia. In some species the conidia are seldom found in long chains
(e.g., Clover Mildew). Spores are
carried by wind to a new host where they may germinate without water being
present. This explains why powdery
mildews thrive under dry conditions.
Spraying with water may even control these fungi. The Ascocarpic Stage has a
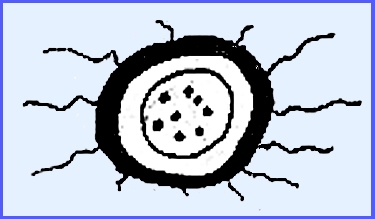
diagrammatic cleistothecia with a well-defined wall of interwoven
hyphae. The color is usually
dark. Appendages grow out from the
outside layer.
A delicate inner layer is present and there is usually only
one ascus inside the ascocarp with less than 8 ascospores. Although there were originally also 8
nuclei the others disintegrate.
Alternatively there may be several asci in a hymenium.
Ascocarps are the overwintering
structures and they are produced in abundance toward the end of the growing
season of plants. The cleistohecium
may explode in the spring followed by an explosion of the asci, which aides
in scattering the spores. Taxonomy is based on appendages
and the number of asci per ascocarp as is shown in Fig 292:
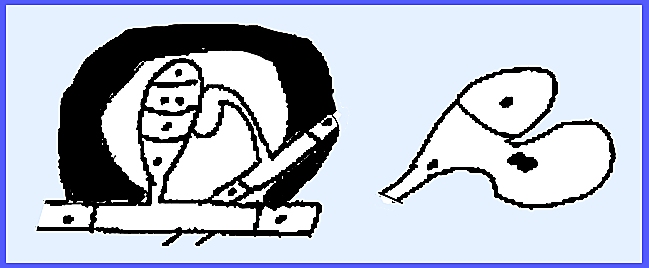
The Genus Spaerotheca
has a Perfect Stage with distinctive characteristics. Many species are heterothallic; and an
ascogonium and antheridium are each produced on separate hyphae. Each possesses a single nucleus.
The male nucleus passes into the
ascogonium through a pore to form the dikaryophase. The ascocarpic wall begins to form by sterile hyphae moving out
from either adjacent cells, the ascogonium or even the antheridium. This process begins before fusion of
nuclei (karyogamy).
As the cell grows it digests the
hyphae that are in the interior of the fructification. These hyphae are the sertoli layer. There are eight nuclei in the ascospore,
but not all may develop into ascospores.
When more than one ascus is
delimited, the ascogonium has previously sent out a system of ascogenous
hyphae.
----------------------------- The families Meliolaceae
and
Capnodiaceae
include the "Sooty Molds." They are not closely related to the
Erysiphaceae and are uncommon in North America, being rather more tropical in
distribution. Information about them
is scanty. The mycelium is largely
superficial but there may be a partial internal mycelium that produces
haustoria. The mycelium is always
dark in color. There are no obligate
parasites and some species even grow as saprophytes on insect honeydew. When on fruit they can disfigure the
surface. Meliolaceae is the most
important family of sooty molds and is one of the most common maladies in
tropical areas. The term "Sooty Molds"
may also be applied to other fungal groups as well. In North-central North America the most prevalent "sooty
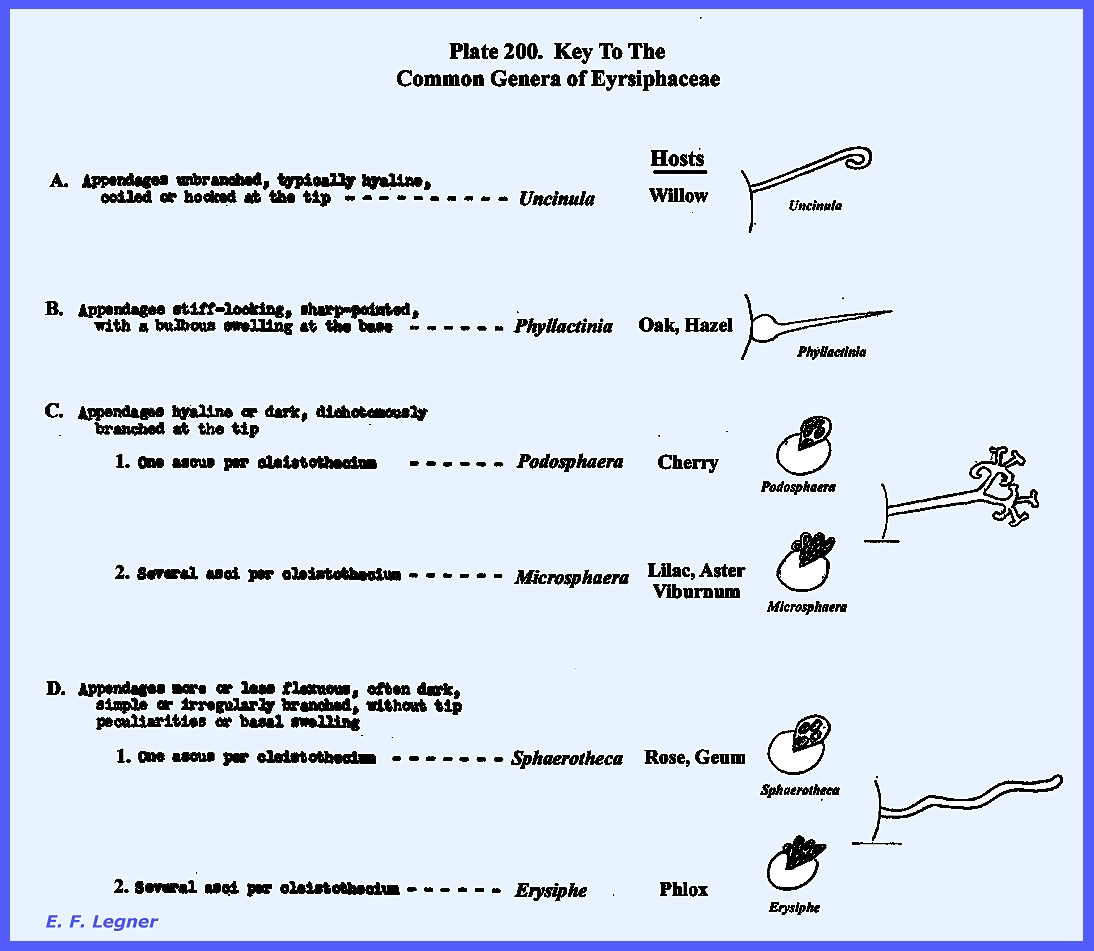
mold" is a Pyrenomycete Apiosporina. Key To The Common Genera of
Erysiphaceae (Plate 200)
----------------------------- Please refer to the
following plates for characteristic structures and Life Cycles in the Erysiphales Ascomycota:
Euascomycetes: Erysiphales Plate
124 = Erysiphaceae: Host cells & mycelia relationships. Plate
125 = Life Cycle -- Sphaerotheca castagnei. Plate
126 = Erysiphaceae: Taxonomic characteristics. Plate 199 = Life
Cycle -- Erysiphales: Erysiphaceae: Spaerotheca sp. Plate 200 = Key to
The Common Genera of Erysiphaceae Plate 201 = Example
Structures & Plant Host Symptoms: Plectomycetes: Erysiphales ----------------------------- In the Sub-Class Euascomycetes, the Series Pyrenomycetes
is a large group with few stable, ordinal characteristics. Their classification has been in a
continuous state of flux with various new alignments being proposed. The changes which have been suggested from
time to time originate from the continuing attempt to make classification of
the fungi as natural as possible.They produce perithecia or perithecia-like
hyphae. A perithecium possesses a
peridium while the "perithecium-like" forms are without such a
peridium. The ostiole is an opening
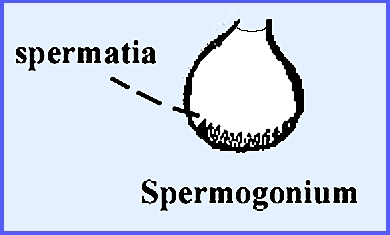
in the perithecium. Among the gametes the female is
the ascogonium and the male the antheridium or spermogonium. The spermogonium produces small,
uninucleate cells at its base, which are termed spermatia. These will generally not germinate but act
rather as non-motile male gametes.
Inside the perithecium there may
occur two kinds of elongated asci.
Most asci are initiated by crozier
formation, and they are always formed at the base of the perithecium as a
hymenium. Sterile hyphae will often
project between the asci on the hymenium.
They may vary in shape and length.
Others may occur just under the ostiole and project into the
opening. The basal structures are
termed paraphyses and the apical ones periphyses. Pseudoparaphyses are sterile hyphae that are joined to the
perithecial wall basally and apically.
Two stages that occur generally in
the Pyrenomycetes are a perfect one that allows for a recombination of
characteristics, and an imperfect one, which does not allow for character
combination. Two orders in the Pyrenomycetes,
Huypoceales and Sphaeriales,
constitute the "core" of the Pyrenomycetes. Previously the genera of both of these
orders have been classified on the basis of the color of their ascocarps, and
so they will be treated here: Hypocreales
= bright-colored ascocarps; soft, waxy, rarely brittle. Sphaeriales = black or brown ascocarps;
leathery and brittle. In the order Hypocreales
the Genus Neocosmospora
is a typical representative. This
is an old order that some authorities have contended should be merged with
the Sphaeriales. Generally the
members of these two groups are basically much alike. They have been segregated primarily on the
basis of what some feel are unimportant characters: principally differences in the color and texture of the
ascocarps. The Sphaerailes as
traditionally defined have ascocarps that are brown to black and carbonaceous
to leathery in texture.
Morphologically similar forms with softer, bright-colored
fructifications have been assigned to the Hypocreales. In both orders the ascocarp is an
ostiolate perithecium and in either the perithecia may be borne singly on the
mycelium or they may be produced in dense clusters, seated upon or imbedded
within a stroma. In the latter case
the color and texture of the stroma are usually similar to that of the
perithecial walls. Whatever the
groupings are there is in the large assemblage of forms producing dark and
bright-colored true perithecia the core group of the Pyrenomycetes. The
perithecium is typically flask-shaped with a well-developed peridium. These are found grouped or distributed
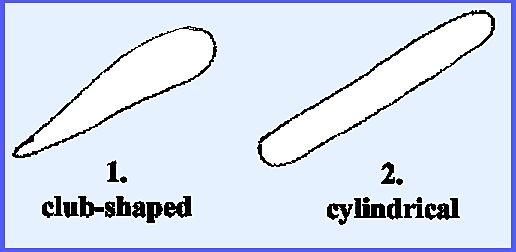
over the mycelium. Elongated asci are
usually cylindrical, which at maturity contain 8 ascospores arranged linearly
inside the asci. At maturity of the
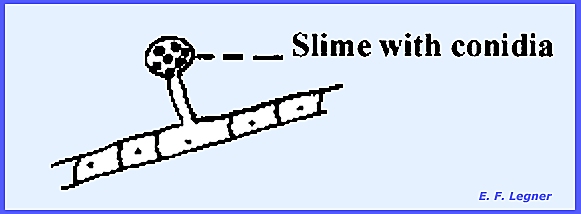
ascospores the neck of the perithecium becomes dark. There is a Cephalosporium Imperfect Stage. Here there are simple conidiophores and
the conidia are suspended in a drop of slime at the apex of the conidiophore:
Perithecia may be borne singly on
the mycelium or in a stroma. The family Nectriaceae is
distinguished by producing their perithecia superficially either on a
well-developed stroma or without a stroma.
Species of the Genus Nectria are
saprophytes on twigs and branches although some are also parasitic. The mycelium is distributed over the host
bark and small patches of stroma form under the bark surface.
The bark ruptures, exposing
orange, cushion-shaped stroma. The Fusarium
Imperfect Stage forms on top of the stroma.
Perithecia develop on the stroma
at the end of summer, with the youngest perithecia appearing at the
apex. They completely cover the
stroma.
Pseudoparaphyses occur in the
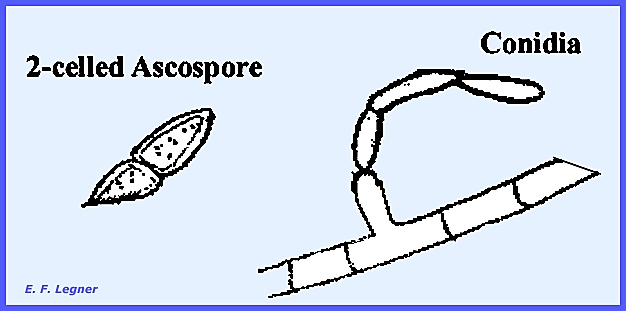
perithecium. Asci contain 8,
two-celled ascospores with a shape similar to Hypomyces. The family Hypocreaceae has
their perithecia either completely or partially buried in a well-developed
stroma. But the existence of
intermediate forms makes separation on this basis difficult. The Genus Hypomyces contain species that are parasites of gill
fungi (Agaricales). The host gills
fail to develop and there is a distortion of tissue. A stroma completely covers the host with a
thin, orange-colored layer.
Perithecia are embedded in the stroma with their necks
projecting. The ascospores are
two-celled. The conidia that are
produced by the Fusarium Imperfect Stage show much variation in size,
shape, number of nuclei, etc.
The family Clavicipitaceae is characterized by
asci that are elongated at maturity and they have a width that approaches
that of the bacteria. A fascile, or
bundle, of filamentous ascospores is produced. In many species crosswalls may come in so that each ascospore
may contain up to 150 cells.
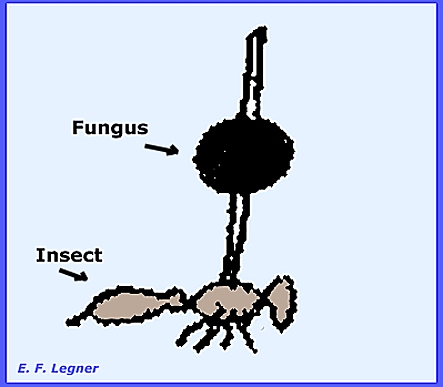
Paraphyses disintegrate before the ascospores mature. The Genus Cordyceps has over 200 species most of them parasitic on
insects. Coremia are usually found in
the Imperfect Stage (Isaria Stage).
Erect, clavate or stalked perithecial stromata also grow out from the
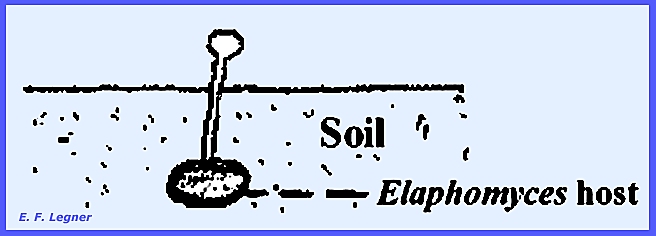
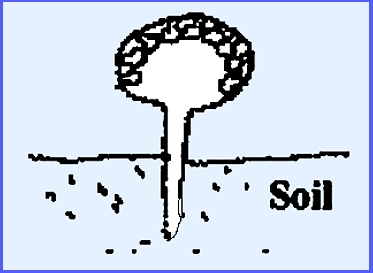
mummified insect. When parasitic on Elaphomyces an easy to locate the
hypogenal ascocarps is to view the brightly colored stromata that appear on
the ground surface.
Perithecia occur on the periphery
of the stroma.
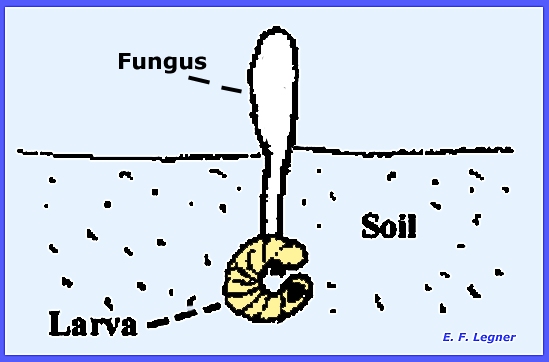
The larvae of the June beetle are
attacked and mummification ensues.
Perithecia develop on the
periphery of an apical club sent up from the host, but not all stroma are
club shaped here.
In the Isaria Imperfect Stage a pyramid of
hyphae with condiiophores branch off.
----------------------------- Claviceps purpurea causes Ergot (or Spur)
Disease of rye, wheat and wild grasses. A mycelium becomes established in the ovary of the host and may
actually replace the ovary. This
mycelium will form a highly convoluted stroma that will be slightly projected
out of the ovary tissue. Conidia are
borne on this "mycelium" and they are mixed with a sticky, sweet,
nectar-like secretion, which is attractive to insect vectors.
The mycelial mat hardens into a
stroma, which takes a spur-like form or a "pseudoparenchymatous
sclerotium." This is
slightly longer than the grain pieces that it assimilates.
These drop to the ground in autumn
and form the overwintering stage. In
the spring these sclerotia will germinate and send out tubes that form a
stroma at their apices, which in turn bears the perithecia. The ascospores may be one-or two-celled.
With regards to the spur produced
as the overwinter stage, a toxic alkaloid is produced within, which causes a
disease of humans and animals called Ergotism. It causes
constriction of the blood vessels and results in a dropping-off of the
extremities. Spurs also produce the
drug called Ergotin,
which has been used in medicine to contract the uterine muscles after
childbirth. ----------------------------- The Genus Epichloe causes "Choke Disease." Some species are also parasitic on
grasses. A mycelium forms a stroma,
which extends around the stem. The
perithecia are embedded in the stroma with their necks extruded. In the Tubercularia Imperfect Stage conidia
are densely packed on a stroma. The Genus Gibberella has pink ascospores and a Fusarium
Imperfect Stage. The mycelium may or
may not produce a stroma. Dark blue
perithecia are produced singly and scattered over the stroma or mycelium. Gibberella
zeae and
closely related species are important pathogens on cereals, inciting various
rots. Gibberella fugihuroi was found to cause rice
plants to grow up to twice the size of normal plants. This took place without a distortion of
tissues. It is believed that this
enlargement is due to both hyperplasia and hypertrophy of the host tissue,
but occurring evenly and without distortion.
The research was developed in Japan but halted during World War
II. A mixture of compounds is
involved, and it is very effective at 1 ppm. on some host species. Certain genetic deficiency may be masked
by applying the chemicals, as in the case of dwarf peas returning to normal
size and bush beans to pole beans. ----------------------------- Please refer to the following plates for characteristic
structures and Life Cycles in the Hypocreales: Ascomycota:
Euascomycetes: Hypocreales Plate 135 = Life Cycle -- Nectria cinnabarina. Plate
136 = Life Cycle -- Claviceps purpurea. Plate
137 = Structures of Claviceps purpurea. Plate 202 = Life
Cycle -- Pyrenomycetes: Hypocreales: Nectria sinnabarina Plate 203 = Life Cycle
-- Pyrenomycetes: Hypocreales: Claviceps purpurea Plate 204 =
Diagnostic Characters: Pyrenomycetes:
Hypocreales: Claviceps, Cordyceps, Epichloe,
Gibberella, Hypomyces, Nectria, Neocosmospora Plate 207 = Example
Structures -- Pyrenomycetes: Hypocreales:
Hypomyces, Nectria,
Neocosmospora Plate 208 = Example
Structures -- Pyrenomycetes: Hypocreales:
Claviceps purpurea, Gibberella
zeae, Cordyceps agariciformis,
C. ophloglossoides & Epichloe
typhina =
= = = = = = = = = = = = = = |























{kind=link}