File: <frungi.htm> <Index to Mycology> Pooled References <Glossary> Site
Description <Navigate to
Home>
|
Mycology: The
Study of Fungi1
Including Some Bacteria and Slime Molds (Contact) Please CLICK on underlined links & included
illustrations for details;
Use Ctrl/F to search for
Subject Matter: (Future taxonomic changes will require
agreement among specialists)
Introduction Mycology is of importance in foreign
explorations for natural enemies of invading plants and insects
especially. There have been some
significant successes in Biological Weed Control and it is important to
recognize if potential arthropod importations may be infected with a
pathogen. The following should
acquaint explorers with a general knowledge of what organisms may be present
as they go about their searches for potential biological control candidates. Sections on Schizomycophyta,
Amoebozoa
and Eumycophyta
follows the classification that prevailed from the latter third of the 20th
Century until the present. The
arrangement of the various subgroups is based on the presumed evolution of the most primitive to the more highly
advanced organisms with previous names of groups being included in
parentheses. Although further
rearrangements are expected as more biological and biochemical data are
forthcoming the presented design should enable identification of major
orders, families and genera. Emphasis has been placed on morphological and
behavioral characteristics, and a simple diagrammatic style is used for most
of the illustrations. A binocular microscope with a 20X magnification is
advisable for students wishing to view living and preserved specimens. Greater detail on a particular group of
fungi may be found by referring to publications listed in the References or through
Internet searches. This is a self-contained database
with a minimum of links outside its limits.
Independent Internet searches are encouraged for greater detail on
particular fungal or bacterial groups, close contact with professional
mycologists is required. Background &
Overview
The first scientific effort to classify
the fungi was made by Anton De Bary in 1860.
He divided the fungi into four groups: Saprophytes
(nutrients derived from dean organic material), Facultative Parasites (able to become parasitic but generally
saprophytic, Facultative Saprophytes
(able to become saprophytic but generally parasitic, and Parasites (only able to survive on a
living host).
Mycology was originally a branch of botany, but fungi are
evolutionarily more closely related to animals than to plants albeit this was
not widely accepted until the late 20th Century. There have been
many schemes developed to classify organisms (see Systems & Kingdoms) and fungi in
particular. Two contemporary proposals
to classify fungi are shown in Table
1 & Table 2. Historically, the bacteria and slime molds were also
included under the broad group "Fungi" until they were separated
into "Kingdoms" of their own (Table
1) (Also See Wikipedia). All of these are alike in one
respect: they lack chlorophyll and
thus cannot make their own food.
They, like animals, depend for their food either directly or
indirectly on green plants. The following sections discuss
diagnostic structures that aid in the identification of the major organism
groups in an arrangement that begins with primitive forms and proceeds to the
more advanced. Included are Bacteria
(Monera, Schizomycophyta), Slime Molds (Amoebozoa), and
the True Fungi
(Eumycophyta) and their
principal Classes: Zygomycota
(zygote fungi), Ascomycota (sac fungi), Basidiomycota (higher fungi), and
Deuteromycota (Fungi Imperfecti). Representative Genera and some species of
major families are included. Of
special interest is that sexual processes that appear throughout these groups
gradually disappear as they ascend the evolutionary ladder. The systematic study of these organisms is
scarcely two hundred years old, but humans have known the manifestations of
this group of organisms for thousands of years. Yet today few realize how intimately our lives are linked with
them. They plan such an important
role in the slow but constant changes taking place around us because of their
ubiquity and their amazingly large numbers.
They are the agents responsible for much of the disintegration of
organic matter, and as such they affect us directly by destroying food and
fiber and other goods that are manufactured from raw materials subject to
their attack. They cause the majority
of plant diseases and man7y diseases of animals and humans. They are the basis of a number of
industrial processes involving fermentation, such as making wines, bread,
beers and even the fermentation of the cacao bean and the preparation of
certain cheese. They are deployed in
the commercial preparation of many organic acids and of some vitamins, and
are responsible for the manufacture of a number of antibiotic drugs, notably
penicillin. Fungi in particular are
both destructive and beneficial to agriculture. On the one hand they do extensive damage to crops by causing
plant disease, while on the other they increase the fertility of the soil by
inducing various changes that eventually result in the release of plant
nutrients in a form available to green plants. Their widespread use as edible food in the form of mushrooms
also should not be overlooked. ============================= The fungi rank prominently in numbers of
species among organisms.
Comparison estimates of some species as of 2020 are noted as follows: Cyanophyta (Cyanobacteria) = 1,800
species Euglenophyta
(Euglenozoa) Flagellate protozoa = 350 species Chlorophyta (Green algae) = 3,250 species Protista (Chrysophyta) (Golden
algae)= 5,225 species Phacophyta (ProtistaBrown algae)
= 1,675 species Rhodophyta (Red algae) = 2,810
species Dinoflagellata (Pyrophyta -- fire algae) = 1,215 species Monera (Bacteria) (Shizomycophyta -- = 2,200 species Protista (Myxomycophyta) = 535
species Eumycophyta (Eumycetes) True
fungi = 142,000 species Bryiophyta (Mosses) = 28,000 Tracheophyta (Vascular plants) =
380,000 (most likely many more species exist) The five main
food sources that are required by fungi are Carbon, Nitrogen, Minor elements,
Vitamins (Thiamin & Biotin) and Oxygen.
The main carbon source is Sucrose, but one group, the Mucorales, is
unable to use it as a source of carbon. Main Groups of Fungi
Zygomycota (Phycomycetes) -- zygote fungi Ascomycota (Ascomycetes) -- sac fungi Basidiomycota (Basidiomycetes) -- higher fungi Deuteromycota (Deuteromycetes or Fungi
Imperfecti) anamorphic fungi General Characteristics of Fungi
The fungi are a group of living organisms that do not possess
chlorophyll. They resemble green
plants as generally they have definite cells walls, they are usually
nonmotile, although they may have motile reproductive cells, and they
reproduce by means of spores. They do
not have stems, roots or leaves, nor dor they have a vascular system as the
more advanced types of plants. Fingi
are usually filamentous and multicellular;
their nuclei can be seen with relative ease; their somatic structures with
few exceptions show little differentiation and practically no division of
labor.
The filaments that make up the body of a fungus elongate by apical
growth (Plate 51). However,
most parts of an organism are capable of growth, and a tiny fragment from
almost any port of the fungus is enough to start a new individual. Reproductive structures are differentiated
from somatic structures and show a variety of forms, which are useful for
identification. Few fungi may be identified
if their reproductive stages are not available. This is becuase with few exceptions the somatic parts of fungi
resemble those of many other fungi.
The fungi obtain their food either by infecting living organisms, by
behaving as parasites, or by attacking dead organic matter. Most fungi, whether normally parasitic or
not, are able to live on dead organic matter, which makes it possible to grow
them on synthetic media. Fungi that
live on dead matter are unable to infect living organisms and are referred to
as obligate saprobes. Those capable
of inciting disease or of living on dea organic matter are referred to as
facultative parasites or facultative saprobes. Those that require living protoplasm are obligate
parasites. Fungi also differ from
most plants in that they require already elaborated food to live and are
incapable of manufacturing their own.
But, if provided with carbohydrates in some form most fungi can
synthesize their own proteins by utilizing inorganic or organic sources of
nitrogen. Many fungi can synthesize
vitamins, which they need to grow and reproduce as do other organisms. Excess food is usually stored in the form
of glycogen or oil.
Fungi vary in their food requirements. Some are omnivorous and can live on anything that contains
organic matter. Other fungi are more
restricted in their diet and a few of the obligate parasites not only require
living protoplasm but are also highly specializes as to the species and even
the variety of host they parasitize.
Enzymes determine what foods are able to be used. The Fungus Vegetative Body
The mycelium is
the entire vegetative body of a single thalus. It is composed of thread-like structures or hyphae. The diameter of a hypha varies between 2
and 50 microns. Branching occurs
behind the tip, there being some degree of apical dominance. Walls in the Zygomycota (Phycomycetes) are
principally of cellulose,
while in the other groups the walls may contain a combination of cellulose
and fungus
chitin. The mycelial type
of thallus is not present in many of the lowest members of fungi, and a few
degenerate forms in the higher fungi also lack it. Occasionally, as in the
Zygomycota, a single hypha will compose the entire mycelium and cross walls
will form at random. The absence of
cross walls is known as coenocytic, and is also characteristic of the
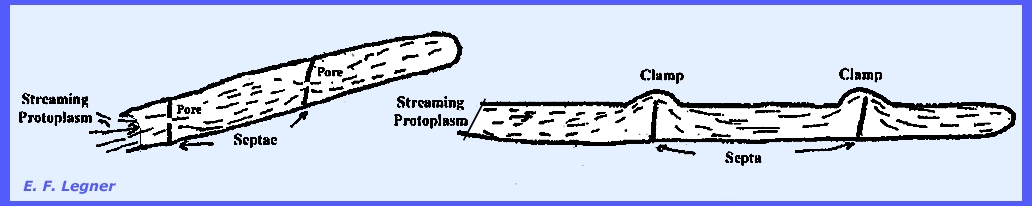
Zygomycota. Septa
are the cross walls and are characteristic of the higher true fungi (Plate 52b). In some species a pore (hole) is left in the septa and protoplasm
is continuous from cell to cell of the hypha. In the Basidiomycota
(Basidiomycetes) a characteristic feature is that a clamp
is formed, and the septa do not reach to the end of the diameter.
All true
fungi (Eumycophyta) have well-defined nuclei. In the coenocytic condition there may be nuclei distributed
throughout the hypha (Plate
52a). When mycelia occur
in the Phycomycota they are characteristically of the coenocytic type. Septations (cross-walls in the hyphae) are
almost entirely lacking.
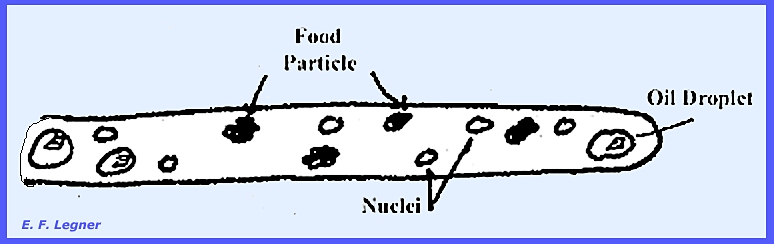
Vacuoles
and food particles and
oil droplets also
are distributed throughout the mycelium.
Again in the coenocytic condition nuclei may or may not (usually not)
exhibit conjugate nuclear dividion.
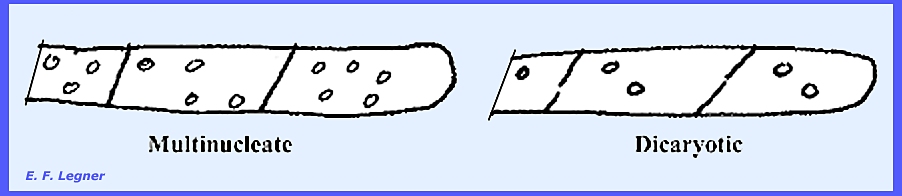
In a septate
mycelium (with or without septal pores),
some species may have many nuclei distributed in one cell (multinucleate). Other species may have two nuclei per cell
(dicaryotic). A dicaryotic cell will usually exhibit conjugate
nuclear division, which is the simultaneous division of the two
nuclei in a dicaryon. This gives rise
to four daughter nuclei. These
generally become separated by a septum into two cells, the sister nuclei
migrating into different daughter cells.
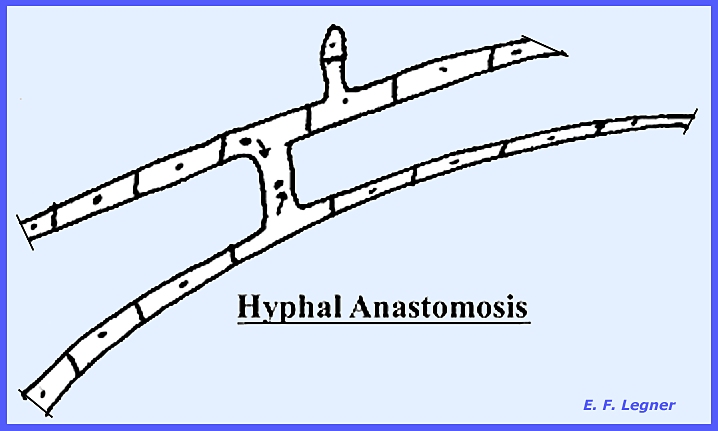
Also, in the septate
condition (septa =
cross wall), the two hyphae will fuse (hyphal anastomosis). This may occur between hyphae on the same
mycelium or on two closely related species.
In this condition the nuclei may migrate over the bridge. Further division may result in daughter
nuclei migrating through septal pores to adjoining cells. This results in a heterocaryotic
effect, which is where there are different nuclei in the
mycelium. It allows for a
recombination of characters. Most
Ascomycota, Basidiomycota and Deuteromycota form mycelia the hyphae of which
are divided by septa.
All hyphae
do not have the same growth rate.
Nevertheless, some forces keep the total margin at an even level
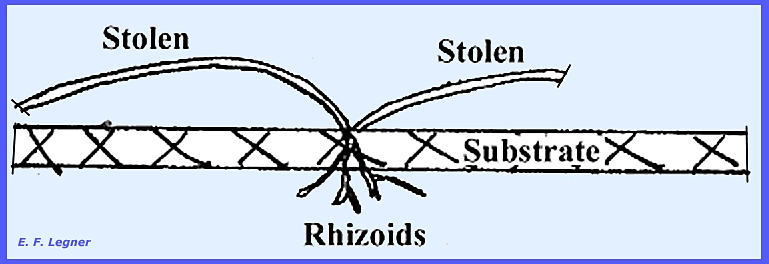
Stolons are
parts of hyphae that skip across the substrate surface. At points of contact with the substrate,
growth is stimulated and hyphae will penetrate the substrate. These penetrating hyphae are then called rhizoids.
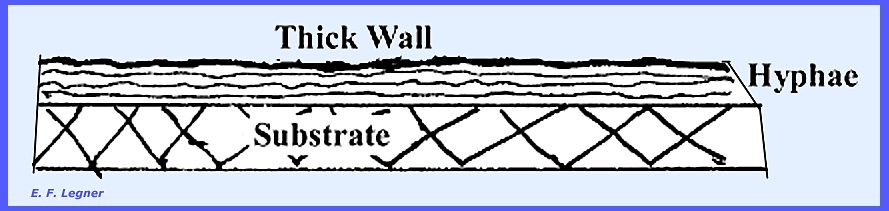
In some Basidiomycota
(Basidiomycetes) a bunch of horizontal hyphae will form a cable over
the substrate (rhizomorphs). This is typified in Armelaria, the
shoe-string fungus. The outer edge of the hyphae forms a thick
cell wall. Its function is believed
to be the transportation of water across dry areas. The cables are usually large enough to be readily viewed
without a microscope and resemble small roots of a seed plant.
Often
the fruiting
bodies will arise from rhizomorphs,
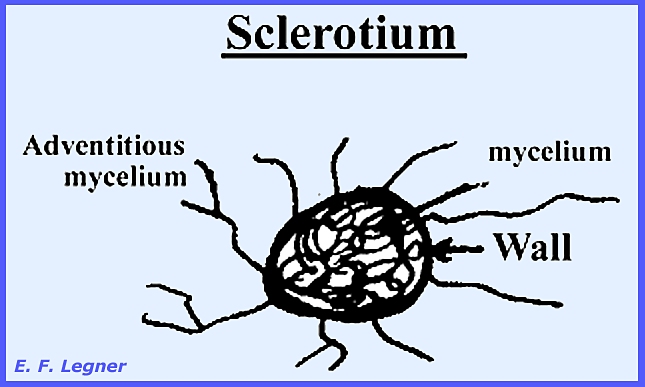
which is particular true of stinkhorn fungi. Sclerotia
(sing. = sclerotium) is a very dense, heavy-packed group of hyphae
surrounded by a thick wall (Plate 56a,b). They are usually found in the higher fungi,
and in certain genera and species they can be of considerable size. The outer hyphae are usually thick-walled
so that the whole structure appears firm and hard. The color is mostly brown or blackish even though the rest of
the mycelium may be white. Sclerotia
may store food and serve as resistant vegetative resting structures when they
occur (Plate 56c,d).
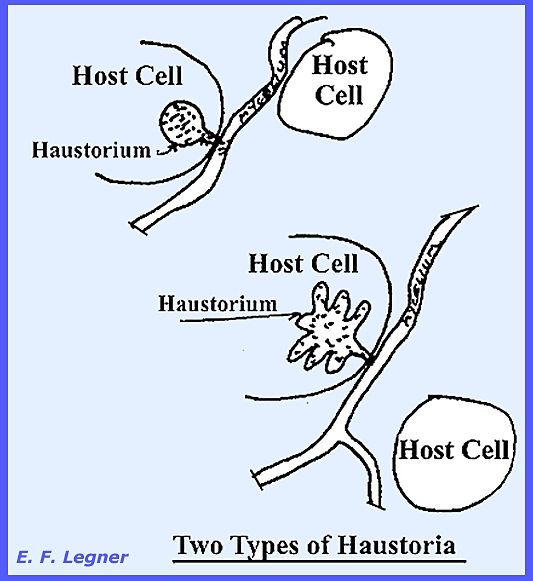
Haustoria
are usually found among the obligate parasites where they occur in the intercellular
hyphae (= a protuberance that dissolves the host cell wall and
develop into the cell (Plate 53). There are various kinds and they serve as
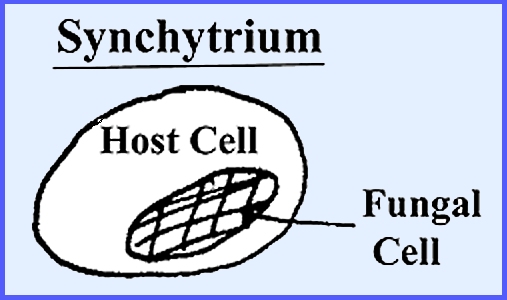
identification characters for certain species. Naturally they do not occur in an intracellular parasite. Some Eumycophyta are not myceliar and are
characterized by a single cell (e.g., yeasts).
[Please
see PLATE 1 and PLATE 2 for additional examples of fungal
vegetative bodies.] Nature &
Reproduction of The Fungi
History
Ancient cultures were well aware
of fungi, but they knew mainly the fleshy kinds. They did not associate the parasitic forms, such as rusts and
mildews, with disease. They were
often amazed at the rapidity of growth.
Theophrastus
(3-4 BCE) believed that fungi were plants without roots, stems and
leaves. The Greeks and Romans formed
the spontaneous generation idea of fungus origins. Pliny (1 AD) proposed that lightening and thunder were
implicated in the rise of fungi. He
observed fairy rings
of fungi,
which are actually the expanding mycelium.
Anton DeBary in 1850 noted that
fungi develop from spores. Franz Unger in
1840 advocated that fungi were associated with disease and were the results of disease. He believed that the morbid sap of the host was
transformed into the fungus. Micheli in Florence, Italy
published Novum
Plantarum Genera in 1729. He described fungi along with other plants in this book, but he
did not believe in spontaneous generation.
He thought that fungi also had seeds (viewed their powdery
spores). In a classic experiment he
used two sterile melons, which he placed under bell jars. He inoculated one and left the other as a
control. Mycelia developed on the
inoculated portion, which he compared with that of the parent. He repeated the experiment several times
and concluded that because of their lightness, fungal spores were in the air
at all times. In a second experiment
he seeded an area in the leaf mat of a forest with non-indigenous species of
mushroom. Later he observed mycelium
and still later the fruiting bodies. Reproduction in the fungi is
varied and sometimes very complex. A sexual
process, or the equivalent, is often involved. However the fungi are noted for the diversity of means they
possess for asexual reproduction. ------------------------------------------- Some fungi employ fragmentation of
hyphae as a means of propagation. The
hyphae break up into their component cells, called oidia, which behave like spores
(Plate 57a). If the cells become enveloped in a thick
wall before they separate from each other or from other hyphal cells
adjoining the, they are called chlamydospores (Plate 57b). Fragmentation may also occur accidentally
by the breaking off of parts of the mycelium through external forces. Such pieces of mycelium under favorable
conditions can start a new individual.
Laboratory propagation is frequently made from mycelial
fragments. Fission can occur through
the simple splitting of a cell into two daughter cells by constriction. This is found among the bacteria
generally, but some fungal yeasts may do this also (Plate 58a). Budding is the asexual
production of a small outgrowth from a parent cell. The bud increases in size while still attached to the parent
cell. It eventually breaks off and
forms a new individual (Plate 58b). Sometimes chains of buds form a short
mycelium. Most yeasts have budding,
but it also occurs in many other fungi at different phases of their life
history or under certain conditions of growth. The commonest method of asexual
reproduction in fungi is by means of spores.
Spores vary in color, size, shape, number of cells and the way that
the spores themselves are borne (Plate 58b) ------------------------------------------- Spores are
small, detachable bodies, with either one or more cells and capable of
germinating (Plate 54). Most fiungi produce these small detachable
bodies, the function of which might be compared to that of seeds in higher
plants. Although there are many spore
types in the fungi, this discussion will stress basically five different
types: Conidia,
Sprangiospores, Zoospores (Planospores), Ascospores and
Basidiospores. Other types include aeciospores,
uredospores,
pycnospores,
etc.
Nevertheless, a number of fungi form more than one type , e.g., both
ascospores and conidiospores), usually at different stages in their
development. The spores may be either
colored or hyaline and exhibit a variety of shapes. They are frequently unicellular but may be two- or more celled. Some fungi bear them on or within a fruiting body,
which consists of a dense aggregation of hyphae. The spore output of some fungi is in the millions or even
billions of spores being produced by a single individual. They are distributed in a variety of ways,
but when they travel by air currents they can be the source of severe allergies as
they are breathed in and begin to germinate on the linings of respiratory
systems in humans and animals. During some stages of the life
history of most fungi the mycelium becomes organized into loosely or
compactly woven tissues as distinguished from the loose hyphae ordinarily
making up the thallus. The general
term plectenchyma is
used to designate all organized fungal tissues. Two types of plectenchyma are
prosenchyma,
which is a loosely woven tissue where the component hyphae lie mostly
parallel to one another and their typically elongated cells are easily
distinguishable; and pseudoparenchyma which consists of
closely packed, generally isodimetric or oval cells that resemble the
parenchyma cells of higher plants. In
this type of tissue the hyphae have lost their individuality and are not
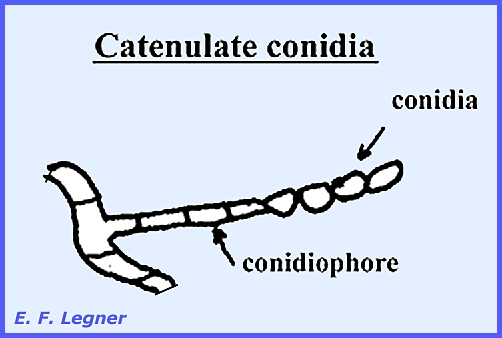
distinguishable as such (Plate 55). Conidia are small, detachable
bodies, either with one or more cells and capable of germinating. Catenulate conidia are
borne in chains. They may become
catenulate by continuous pinching-off of the end of the conidiophore, or the first
conidium may divide giving rise to the second, and so on:
Sporangiospores
are common in the Phycomycota:
Zoospores (Planospores)
are characteristic of aquatic fungi:
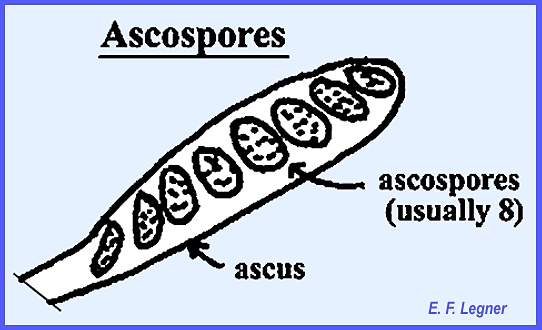
Ascospores are
characteristic of the Ascomycota, although these also exhibit other spore
types.
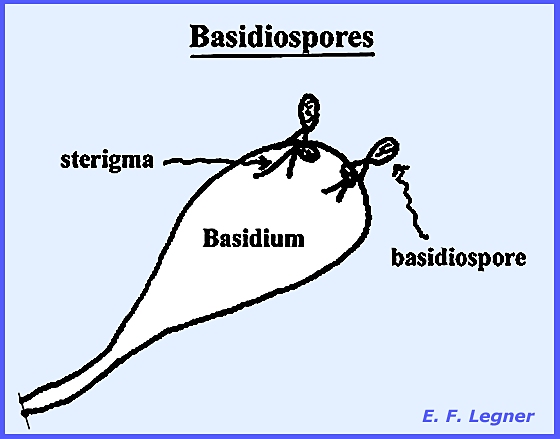
Basidiospores are
characateristic of the Basidiomycota, although these also exhibit other spore
types:
All the noted spores are walled structures except the
zoospores Provost in 1807 first observed a
spore germinate from one fungus species.
In the process of germination a spore must have a suitable environment
(water taken up). The wall becomes thin
in one or more places after water has been taken in.
In double-walled spores, the outer
wall cracks on germination.
Viability may be either long or
short. Some spores are not durable at
high altitudes (high ultra-violet rays cause lethal mutagens). Sometimes simply the presence of an
element, e.g., Boron, will stimulate germination. In multicellular spores each cell can give rise to a mycelium.
------------------------------------------- In 1952 Alexopoulos gave a
detailed narrative of Sexual Reproduction in the fungi, which holds true into
the 21st Century, and the following description is derived therefrom [Alexopoulos,
C. J. 1952. Introductory Mycology.
John Wiley & Sons, NY. 482
p.].
The most common
methods where compatible nuclei are brought together (Plasmogamy)
are:
2. Gametangial contact. In a large number of fungi, the gametes of
the male or of both the male and the female gametangia have been reduced to
undifferentiated protoplasts consisting chiefly of a nucleus. Such gametes
are never released from the gametangia to the outside, but are transferred
directly from one gametangium into the other. In this method, two gametangia
of opposite sex come in contact, and one or more gamete nuclei migrate from
the male to the female. In no case do the gametangia actually fuse or in any
way lose their identity during the sexual act. The male nuclei, in some
species, enter the female gametangium through a pore developed by the
dissolution of the gametangial walls at the point of contact; in other
species, an especially developed fertilization tube serves as a passage for
the male nuclei (Plate 60). After the
passage of the nuclei has been accomplished the oogonium continues its
development in various ways, and the antheridium eventually disintegrates.
a. Passage of the
contents of one gametangium into the other through a pore developed in the
gametangial walls at the point of contact.
This method is typical of some holocarpic forms in which the entire
thallus acts as a gametangium, the male thallus attaching itself to and
emptying its entire content into the female thallus (Plate 78f).
b. Direct fusion of the
two gametangial cells into one. This takes place by the dissolution of the contacting
walls of the two gametangia, resulting in a common cell in which the two
protoplasts mix (Plate 61, 111g, 112g). 4. Spermatization. Some
fungi bear numerous, minute, uninucleate, spore-like, male structures termed
spermatia that h are produced in various ways. The spermatia are carried by insects, wind, water, or (in some
other way, to the female gametangia or to special receptive hyphae, or even
to somatic hyphae, to which they become attached. A pore develops at the
point of contact, and the contents of the spermatium pass into the particular
receptive structure that serves as the female organ (Plate 62)
Sexual
Compatibility Sexual compatibility. Although this phenomenon
is certainly related to sex because it affects sexual reproduction,
compatibility should not be confused with sex. There are, for example, many fungi that produce clearly
distinguishable male and female sex organs on the same thallus but in which,
nevertheless, single individuals are sexually self-sterile because their male organs are incompatible with their female organs and no
plasmogamy can take place. On the
basis of sex, most fungi may be classified into three categories: 1. Hermaphroditic, in which each thallus bears both male
and female organs.
morphologically
indistinguishable as male or female. Fungi in
the above sex categories belong to one or the other of the following two
groups on the basis of compatibility: 1. Those in which every thallus is
sexually self-fertile, and can therefore reproduce sexually by itself without
the aid of another thallus.
Life
Cycle in Fungi
------------------------------------------- Please view the following for additional examples of Fungal
Structures & Reproduction: Plate 1 = Fungal Vegetative Body-1 Plate 2 = Fungal Vegetative Body-2 Plate 3 = Examples of Fungus Spores Plate
51 = Successive growth stages of hypha: Gelasinospora autosteira. Plate
52 = Somatic hyphae. Plate
53 = Three types of haustoria. Plate
54 = Two stages in spore germination. Plate
55 = Fungal tissues: Parenchyma & Pseudoparenchyma. Plate
56 = Stroma & sclerotium: Daldinia sp. & Claviceps
purpurea Plate
57 = Asexual reproduction: Fragmenting
hypha: Collybia conigena &
Fusarium sp. Plate
58 = Asexual reproduction: Budding Plate 58b = Various types of fungal spores. Plate
59 = Sexual reproduction: Planogametic copulation: Catenaria sp.,
Allomyces arbuscula & Monoblepharella
taylori. Plate
60 = Sexual reproduction: Plasmogamy thru' gametangial contact in Pythium
aphanidermatum. Plate
61 = Sexual reproduction: Plasmogamy thru' gametangial copulation in
Sporodinia garndis. Plate
62 = Sexual reproduction: Plasmogamy by spermatization in Pleurage
anserina. Plate
63 = Sexual reproduction: Plasmogamy thru' somatogamy in Peniophora
sambuci. |
|||
|
|
|
|||