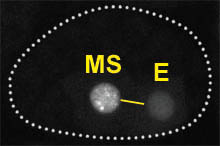
The EMS cell,
formed at the 4-cell stage, is a mesendoderm progenitor: EMS
divides to produce an anterior cell, MS, and
a posterior cell, E. The E cell generates
the entire gut (endoderm) while MS generates mesodermal cells
(including muscle and part of the feeding organ, the pharynx).
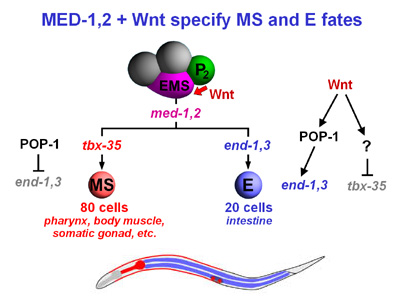
We have shown that both cells are specified by the genes med-1,2.
How do the
same genes specify two very different fates? The MS and
E cells are made different from each other at the 4-cell stage.
The
neighbor of EMS, called P2,
polarizes EMS so that the daughter cell closest to P2 becomes
the E cell. The P2-EMS
signal uses components of the conserved Wnt signaling
pathway, which is important for many developmental processes (including
cancer in humans). The endpoint of Wnt signaling in the MS/E decision
is the differential activity of a regulator called POP-1. In
the MS cell, POP-1 represses the endoderm fate by directly repressing
the E-specifying genes end-1 and end-3. In the
E cell, this activity of POP-1 is blocked.
GFP::POP-1 localization in vivo
We
have shown that the asymmetric activity of POP-1 results from
the movement of POP-1 from the nucleus to the cytoplasm. This
results in high nuclear levels of POP-1 in the anterior cell,
MS, and low nuclear levels in E. POP-1
movie
Using an in
vivo assay that allows visualization of protein-DNA
interactions, we found that GFP::POP-1 directly binds to
the end-1,3 genes
in living embryos.
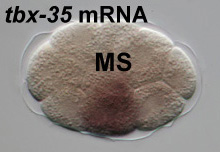
localization of tbx-35 mRNA in situ
Our
recent work has shown that in MS, MED-1,2 activate
the regulator tbx-35. Embryos lacking tbx-35 arrest
without most of the tissues normally made by MS. Shown at left
is the localization of tbx-35 mRNA in an early embryo,
using an in situ hybridization protocol developed in
our laboratory.
In the absence of POP-1, tbx-35 continues
to be activated in the MS cell, suggesting that an additional
nuclear Wnt coregulator
specifies MS-E differences.
We have found that while the MED-1,2 regulators structurally related
to the broad GATA class of transcription factors, they
bind a
non-canonical
target
site. As med-like genes are found only in Caenorhabditis species,
we hypothesize that this subclass of regulators
evolved very recently. We have an ongoing collaboration with
the laboratory
of Joel
Mackay, University of Sydney, Australia to study the
solution
structure
of the MED-1 DNA-binding domain. (See MED-1 Structure page.)
More recently we have found that ceh-51, a target of TBX-35 and at least one other factor, provides MS-specifying functions in tbx-35 mutant embryos. As a result of this, double mutant ceh-51; tbx-35 embryos have a strong MS defect that is similar to the MS defect in med-1,2 mutants. Part of this work was performed in collaboration with Paul Sternberg (Caltech).
We are currently trying to further elucidate the genes that function in MS specification downstream of TBX-35 and CEH-51. Other current projects
include examination
of expression patterns of other embryonic
genes,
and evaluation of differences in the endomesoderm gene network
in other nematode
species.