File:
<pollinat.htm> <Index
to Pollination> Site Description <Navigate
to Home>
|
An
Introduction to Insect Pollination &
Bee Keeping (Contact) Insects
especially are of enormous importance in the pollinations of many
agriculturally important crops. Although
gravity, wind, water, mollusks, birds, bats and humans are agents of
pollination, it is often possible to manipulate insects in their performance
on high value cropping systems. This
section discusses the processes involved in plant reproduction and
pollination with emphasis on agriculture.
The kinds and numbers of insects of importance in these processes are
detailed. Links (blue &
underlined) are posted that refer to subject matter within this database;
photos and illustrations of the insects involved may be viewed by clicking on
underlined insect groups in the text and by referring to <Beneficials> or <Families>. Appreciation is extended to George E.
Bohart, Donald L. Davis, Department of Entomology, Utah State University and
the U. S. Dept. of Agriculture, Logan, Utah (see USDA) for inspiring the creation of this section. Citations CONTENTS Background and
Overview
Pollination of
plants may involve two basic procedures:
Self-pollination and Cross-pollination. In Self-pollination the transfer of pollen is from the anther
to stigma of the same plant or to another plant having the same genetic
composition. With an identical genome
the two would either belong to a single clone or to members of an entirely
Homozygous variety. Clones or clonal
varieties are composed of a series of plants that have been propagated
vegetatively from a single plant.
This type of reproduction does not cause any change in the genetic
makeup of the offspring. A completely
homozygous plant possesses sex cells where the two parental sets of
chromosomes are identical. Thus when
they undergo reduction division, the products of the division are
identical. When such plants are
self-fertilized, the offspring will be equal genetically and pollen transfer
between them does not introduce any new characteristics. Two types of Self-pollination are
Auto Self-Pollination and Indirect Self-Pollination. In the Auto-type there is no external
agent of transfer. Pollen is
transferred within one flower or between adjacent flowers. In the Indirect-type the pollen transfer
involves an external agent. When such
pollination occurs within one flower, insects are usually involved. When it is between flowers on the same
plant, both insects and gravity are involved. When it is between plants that are completely homozygous or
from a single clone, both insects and wind are the main agents of
transfer. For example, in some
varieties of flax that are nearly homozygous, seed from fields of such
varieties is especially uniform even where much pollen interchange has
occurred from plant to plant. Another
example is the pollination between plants in a vineyard of a single variety
of European grapes that were propagated by stem cuttings. Three type of Cross-pollination
are Intra-varietal, Inter-varietal and Interspecific or Intergeneric. In the Intra-varietal type pollen transfer
is between two plants of a single variety.
An example is the pollination between plants of cauliflower (Brassica
oleracea var.). For the
Inter-varietal type, pollen is transferred between two varieties that differ
widely in their genetic makeup. An
example is the pollination between cauliflower and cabbage (two varieties of Brassica
oleracea). In the Interspecific
or Intergeneric type, pollen transfer is between separate species or
genera. An example is the pollination
between cauliflower and turnip (two distinct species of Brassica). Pollination and
Plant Breeding. Pollen is basically a spore that has
been produced asexually. It
germinates on contact with the stigma of a flower and grows as a pollen tube
through the style to the embryo sac where it discharges two nuclei. One nucleus unites with the egg cell of an
ovule and results in fertilization.
The fertilized egg develops into a mature plant. The other nucleus from the pollen tube
unites with the polar bodies to form the endosperm nucleus in the same ovule. The seed endosperm, or nutritive tissue
like yolk in an animal egg, develops from this union. It then dies early in the development of
the young seed or seedling which drains it of nutriment. Sperm cells and egg cells mature by
dividing the number of their chromosomes in half. Then when egg and sperm unite, the original number of chromosomes
characteristic of the cells in the plant is restored. Inheritable traits of the parents of both
the egg and the sperm cells are now combined in the developing embryo. If these parents should be one and
the same (i.e., as a result of self-pollination) no new genetic traits will
be introduced. However, because the
original ones were independently segregated during reduction division, they
can recombine in different patterns so that the plants that result from
self-fertilization may differ somewhat from their parents. Long sustained self-fertilization combined
with artificial selection of one type will eliminate these variations so that
in time the plants can be considered “homozygous” and will breed true when
self-fertilized. That is, unless an
entirely new inheritable variation or “mutation” occurs in the sex cells. Sometimes the attribute of “breeding true” for a desirable type is
desirable for plant breeders, but most of the time the close inbreeding that
is necessary to bring it about results in a loss of vigor or partial
sterility or both. Many plants that
are self-sterile still produce few or no offspring when self-fertilized. An example is alfalfa that produces few
seeds when self-pollinated. Although
relatively self-fertile varieties of alfalfa exist, they are poor in growth
and reproductivity. When plants are cross-fertilized,
the traits of both parents are present in different combinations among the
offspring. Plants from such unions
remain variable and certain individuals with undesirable characteristics may
appear. Nevertheless, vigor and
reproductivity remain high. In plant breeding there is an
effort to achieve both uniformity of desirable traits and continued
vigor. Several methods may be employed
as follows: Sometimes a desirable type of
plant shows up that will self-fertilize without losing much vigor or
reproductivity. If it can also be
easily self-pollinated it can become a standard variety. Examples are in most varieties of wheat. Close inbreeding can segregate at
times desirable traits in two stocks.
These two stocks, weak though they may be, might then be crossed so
that their offspring will again be vigorous and retain the desirable
characteristics of the inbred parents.
Following several generations the crossed stock may again become
variable and the desirable characteristics lost. But this in turn can be prevented by asexual propagation (stem
cuttings, root pieces, bulbs, etc.).
Here the genetics do not change and the desirable traits are
maintained. Some hybrid varieties of
grape are good examples. Also some
plants cannot be propagated vegetatively in practicality. Preserving the inbred lines for breeding
purposes may also prevent it and producing crossed seed from them for use in
one planting only. Examples are the
production of hybrid maize. Uniformity may also be attained by
vegetative propagation of cross-fertilized plants that have been selected but
not inbred for desirable characteristics.
Apples, for example, are vegetatively propagated from seedlings that
were observed to have certain useful traits.
But the seeds of these seedlings will usually be worthless because of
their mixed “heterozygous” inheritance. Controlled crossing may also be
deployed to create uniformity. Here
undesirable plants re rejected. Many
undesirable traits can be eliminated or reduced in this way and at the same
time vigor is retained. Fields used
for seed production of varieties produced in this way must be isolated from
wind or insect transported pollen of foreign varieties that could reintroduce
undesirable traits. An example is
Ranger alfalfa.
CLICK to enlarge
General Botanical Terminology
Asexual reproduction = New
plants formed without the union of sperm and egg cells (as in rhizomes,
tubers, grafts, stolens, etc.) Also known as vegetative propagation. Dioecious plants = These produce
only pollen or only embryo sacs (the sexes occur on separate plants). Fertilization = The union of sperm
cells from pollen grains with egg cells in the embryo sac. Flowers = Structures
on the plant the maintain pollen for the production of sperm cells and embryo
sacs for the production of egg cells. Imperfect flowers = Flowers that
produce only pollen or only embryo sacs.
The sexes occur in separate flowers.
Occasionally a “perfect” flower
will also have one sex sterile or aborted (pseudoperfect). Monecious plants = These possess
imperfect flowers of both sexes on the same individual plant. Perfect flowers = Flowers that produce
both pollen and embryo sacs. Both
sexes occur in the same flower. Pollination = The transfer of pollen from
the pollen-bearing organ, or “anther”, to the receptive surface of the female organ or “stigma.” Reproduction = The formation of new
plants Sexual reproduction = The formation of
new plants from fertilized egg cells.
This is equal to the typical flower- seed-new plant sequence. ---------------------------------------------------------------- Fruit Formation
Pollination
and subsequent fertilization are usually necessary for the formation of a
plant embryo, its adjoining nutritive matter and the protective coats that
together compose the seed.
Fertilization is also usually required to provide a stimulus for the
development of fruit around the seed or seeds. The nature of the stimulus probably involves hormones as shown
in tomato hormones that have been successfully used to provide a substitute
stimulus and have resulted in the development of seedless fruit without
pollination (= parthenocarpic fruit formation). Fertilization of each
ovule in the ovary has been shown to stimulate fruit development in the area
adjacent to it so that each ovule must be fertilized for maximum fruit
development. Incomplete pollination
in plants with more than one ovule usually results in the formation of stunted
or deformed fruit. Apples and
strawberries may be stunted and malformed for this reason. Pollinating Agents
Both surface and
subsurface water may disseminate pollen.
Gravity and wind, sometimes aided by insects, may play a role. Various animals such as bats, birds,
mollusks, insects and humans are frequent pollinators. Humans may be involved by hand
pollination, in breeding efforts, where the natural sources of pollen are
inadequate as in apple orchards, and where natural agents of pollination are
scarce. An example is vanilla which
must be hand pollinated because no pollinators exist in most of the areas
where it is commercially grown. Floral characters that favor
pollination by insects are a conspicuousness of flowers and inflorescences, a
distinct odor, the presence of nectar and a sticky or large pollen grain. Those characters favoring
pollination by wind are an abundant, dry, lightweight pollen, mechanisms for
rapid dehiscence of pollen, anthers exposed to the wind and a feathery
stigma. Cross pollination is favored in
imperfect flowers (including psuedoperfect), dioecious plants, dichogamy
(anthers and stigma maturing at different times), where the stigma is in such
a position that the anther or its pollen cannot touch it, and by the
prepotency of foreign pollen. Characteristics favoring
Self-pollination are perfect flowers, flowers that do not open
(cleistogamous), homogamy (anthers and stigma mature at the same time),
flowers in which the receptive portion of the stigma is easily touched by
anthers or pollen that is shed there from.
In the latter case the parts may be on the same plane and close
together or the stigma is below the anthers, the stigma may grow up through a
ring of dehiscing anthers, the flower may close at night, bringing the parts
close together, or the lobes of the stigma may recoil and contact pollen shed
from the anthers. Self-pollination
may also be favored by self-fertility and a lack of pre-potency of foreign
pollen. Floral characters that favor
cross-pollination at one time and self-pollination at another include
incomplete dichogamy (cross-pollination favored at first but self-pollination
possible later as the parts coincide in maturity), flowers that mature upon
opening at which time cross-pollination is favored, then close at which time
self-pollination is favored. Also
flowers assuming upright positions at day or early in the cycle (the stigma
is above the anthers) and pendant position later (the anthers are above the
stigma). Flowers in which the lobes
of the stigma recoil at the end of the day and contact anthers or pollen
caught in special hairs on the pistil beneath the stigma tend to alternate
cross and self-pollination. Finally,
plants that bear imperfect flowers in the early part of the flowering cycle,
and later they bear perfect flowers. Floral characters that favor
particular types of insects or other animal pollinators are those with a
light or dark color, which are dull or bright, greenish, white or yellow, red
or blue or variegated. They are also
favored if they bear quantities of nectar, a scent, abundant available pollen
and peculiar shapes. Attractive
shapes include size, regular or irregular, depth and breadth of the corolla
tube, special explosive or retractable mechanisms that can be triggered by
the pollinator, special landing structures, the position of the flower, the
structures to exclude certain insects, and special trapping mechanisms to
hold insects. Certain types of
inflorescences, such as heads, racemes, catkins and panicles, are also
attractive. Insect-Pollinated
Flowers Classified According to Insect
Adaptation
Flowers such as
rose, poppy, elderberry and potato, have no nectar but may be scented, they
are generally conspicuous, simple, regular, with pollen freely exposed and
usually abundant. A great variety of
color types are included. Insects
involved are usually Syrphidae flies, soldier flies and pollen feeding beetles.
Many pollen gathering bees, including honeybees, usually frequent these flowers. They are generally unattractive to Colletiidae bees, male bees, bee flies, moths, butterflies and
hummingbirds. Flowers With
Exposed Nectar (Group II).
Maple,
carrot, some elderberries, Euphorbia, poison oak, grapes and saxifrage
flowers are included here. Their
sparce nectar is freely exposed as droplets, the flowers are simple, open and
regular, and the inflorescence is usually inconspicuous and
greenish-white. They attract many
kinds of wasps and short-tongued flies and bees. They are not very attractive to long-tongued bees or flies and Lepidoptera, but some are attractive to honeybees. Flowers With Partly Concealed Nectar (Group III). Examples are
strawberry, cactus, raspberry, stone fruits, many cruciferous species and
buttercups. Here the nectar is partly
concealed by numerous stamens or hairs or overlapping petals. The flowers are usually completely open only
in sunshine and may be moderately to quite conspicuous. White and yellow colors predominate, but
pink can be common. Attracted insects
are Syrphidae flies and short-tonged bees. Some Rosaceae are
also attractive to long-tonged
bees and honeybees. Sawflies are common on many species in Group III, and some beetles and
butterflies may also be attracted. Flowers With Concealed
Nectar
(but not sexual organs
or deep narrow corollas) (Group IV). Currant,
onion, orange, mallow and blueberry are included here. The nectar is completely hidden in pouches
or by hair tufts. The flowers usually
have corolla tubes and may be somewhat irregular. They are generally conspicuous flowers with blue, red or violet
predominating. Long-tongued bees and honeybees are attracted. Also
some short-tonged bees, bee flies, long-tongued wasps, Lepidoptera. Rarely attracted are
most wasps and short-tongued flies and beetles. Social Flowers With Completely
Concealed Nectar (Group V). The Compositae such as dandelion,
aster, sunflower and Scabiosa are included. The nectar is hidden in narrow but not deep corolla tubes, but
access to nectar is blocked by the stigma and the cone of stamens. Pollen is very abundant. The inflorescence is conspicuous because
of the grouping of flowers into heads.
The color groups white and yellow, and red and blue, are
attractive. This group is very
attractive to short and long-tongued
bees, many butterflies and
polleniferous beetles and Syrphidae flies. Insects that visit white and yellow
flowers in this group are akin to those visiting flowers with partly
concealed nectar, while those visiting red, blue and purple flowers are akin
to those visiting flowers with concealed nectar. Hymenoptera Flowers (Group VI). Violets, legumes, sages, mints,
monkshood, Delphinium, iris and some lilies are included here. The nectar is concealed in bilaterally
symmetrical flowers with slightly long corolla tubes closed at the
throat. The sexual organs are usually
partially concealed by modified petals that require operation of a special
mechanism to expose them. They are
usually positioned horizontally, with special landing structures for the
pollinator. These flowers are visited
primarily by medium to long-tongued
bees that can operate the
mechanisms to get at the pollen and nectar.
Lepidoptera
that visit these flowers generally do not operate the mechanism exposing
pollen so they do not accomplish pollination. They are visited in the same manner by long-tongued Conopidae
flies and bee flies. Many have such
deep nectarines as to be accessible only to bumble bees and a few other
insects. Others have tough tripping
mechanisms that require large, powerful bees for pollination. Other bees may bite holes in the corollas
to rob the nectar without pollinating.
One group of Hymenoptera flower might be called “wasp flower.” It has a ventral pouch filled with nectar
and a dull red color. Lepidoptera Flowers (Group VII). This group includes such species
as tobacco, trumpet flowers, honeysuckle, croc gentian, many orchids and some
lilies. The flowers bear nectar at
the base of long, narrow corolla tubes and spurs. They are rather large and conspicuous with a strong scent. Mainly Lepidoptera pollinate these,
but long-tongued Hymenoptera may frequent some species. In tropical areas stingless bees are able
to crawl into the slender corollas and spurs. Long-tongued bee flies may also use them. Hummingbirds and honey birds are also
important pollinators in tropical regions.
Within the Lepidoptera butterfly and moth flowers differ. Butterfly flowers have variable colors and
they usually open and are fragrant during daytime. On the other hand, moth flowers usually open and are fragrant
only at night. They are generally
white or pale colored. Special Types of
Flowers (Group VIII). Nauseous flowers that are
attractive to flies include some umbellifera, calla lilies, skunk cabbage and
many types of saxifrage. They may
give off odors of feces, carrion or ammonia.
They are especially attractive to filth flies, dung beetles and
others. Pitfall flowers are also often
nauseous. Included are
Jack-in-the-pulpit, pitcher plants and Dutchman’s pipe. They capture flies, holding them until
they become covered with pollen, after which they are released before the
stigma is receptive. Pinch-trap flowers include the
milkweeds and some orchids. The
pollen born on “pollenia” fastens onto visitors and are later pulled off in
stigmatic grooves of the pistil.
These are attractive to flies, bees and wasps. Syrphid fly flowers include Veratrum
and Veronica. The flowers bear
radiating streaks that lead to small, definite centers. Two long stamens are able to dehisce on
the back of the syrphid fly when grasped at the base. Only syrphids are able to accomplish this. Small insect flowers include some
aquatic species and euphorbias and figs.
There is an array of minute flowers that are attractive to tiny
insects. The flowers may be clustered
in a hollow receptacle (as in the fig) with an opening to the inflorescence
that is just large enough to accommodate the tiny insect. Importance of
Pollination to Agriculture
Insects in their pollination
activities have a direct impact on the evolution of flora and fauna. It is believed that angiosperm plants and
the more highly evolved insects evolved together. Primitive flowering plants are all insect pollinated. Therefore, grasses and all other
angiosperms arose from plants dependent upon insects. Some beetles, most Hymenoptera, many Diptera
and almost all Lepidoptera are dependent upon materials provided by flowers. Without angiosperms the evolution of
mammals would certainly have been different.
Rodents, herbivores and primates are especially dependent upon the
products of flowering plants. Thus,
angiosperms were a required forerunner to the stocks, which gave rise to
humans, and insect pollination was necessary to the development of
angiosperms. There would be grave consequences
for the flora and fauna were pollinating insects to disappear or cease
pollinating. Many types of plants
would most likely perish eventually because in time they would be dependent
on insect pollination for competitive reproduction. These would embrace by far most of the angiosperms. Certain elements of flora would rapidly
perish. Plants that are usually
propagated by seed are dependent upon insects for adequate pollination. Included here would probably be over half
of the existing species. Plants that
usually propagate asexually could probably survive for many seasons or
generations. But asexual propagants
are very limited in powers of dissemination and those species would have a
fixed genetics incapable of adjusting to changes, which would be expected to
be rapid under such conditions.
Self-fertile plants that are capable of auto-self pollination might be
able to persist longer. However, most
of these are dependent upon occasional crossing in order to retain
vigor. All would require some
crossing in order to retain the genetic plasticity necessary to adjust to
changing environmental conditions. Some plants might survive
indefinitely without insect pollinators and some might increase in the
absence of normal competition. These
include many nut-bearing trees, grasses, all conifers, and various other wind
pollinated plants such as poplars, birches, elms, alders, etc. Even so, many grasses and other plants
most certainly depend upon the surrounding flora for their survival. Those plants that are produced as crops by
humans and propagated by asexual means might also be unaffected. Breeding for disease resistance, for
example, could be done with hand pollination. Nevertheless, there are many
consequences of a drastic reduction and elimination of most floras. These include the loss of plants with
nitrifying bacteria, soil erosion, a drastic curtailment of the human diet,
loss in forage values for livestock, loss of many kinds of animals, loss of
most kinds of wild flowers, and a general upset in the balance of nature,
with unpredictable results. Advanced agriculture manages the
production of products that require pollination, which are primarily fruits
and seeds. Seeds are used for general
plant propagation and for bedded plants.
Some plants like papaya require occasional seeding; alfalfa is seeded
every few years and spinach is seeded annually. Alfalfa and forage grasses often require a large amount of
seed, while tomatoes and melons need little seeding. Plant breeding by crossing, selfing and
selecting is done with pollination and planting with seeds. Plant products that are consumed directly
include cereals, beans, nuts, oils, fruits, preserves and many
vegetables. Seeds such as grains,
oilcake and peanuts are also used for livestock feed. Many seeds are used as medicines, spices
and flavorings. Seeds, fruit oils and
seed fibers are deployed in industry for soaps, paints, plastics, explosives,
alcohol and textiles. Common
Agricultural Crops Requiring or Benefiting From Insect Pollination (Medicinals and Ornamentals Excluded)
Insect Pollinators Other Than
Hymenoptera
There are more insect species than
all other animals and plants combined, the total number estimated to be over
two million as of 2010. Joined
appendages and an external skeleton characterize insects as part of the
Arthropoda. Included are spiders,
crustaceans, centipedes and scorpions. Insects are classified into 28 major orders,
but seven comprise most of the species.
These are, in order of increasing specialization and importance as
pollinators, the Orthoptera
(cockroaches, grasshoppers, crickets, walking sticks, praying mantis), Hemiptera
(true bugs, cicadas, leafhoppers, scale insects, aphids), Thysanoptera
(thrips), Coleoptera
(beetles), Diptera
(flies, gnats, mosquitoes), Lepidoptera (moths and
butterflies), and Hymenoptera
(ants, wasps, bees, sawflies, Ichneumon flies and chalcid flies). For the most part the Orthoptera of no
importance as pollinators. Only a few Hemiptera of value are Anthocoridae (minute pirate
bugs), Phymatidae
(ambush bugs) and Reduviidae
(kissing bugs). The Anthocoride prey
on thrips in flowers; a few Reduviidae prey on bees in flowers and most
Phymatidae prey on bees and flies in flowers. Anthocoridae are found in almost any flowers that are visited
by thrips. Phymartids and reduvids
are found primarily on Compositae and flowers that are grouped into tight
heads. Except for a few flower-inhabiting
forms, the Coleoptera are not
as important pollinators as the Diptera, Lepidoptera and Hymenoptera. There are nine families of Coleoptera that
are at times involved in the pollination of flowers. Most species of Cantharidae,
the leather-winged beetles, that are predaceous as larvae occasionally
pollinate.. Polleniferous species are
also predaceous as adults. The
majority of Meloidae,
or blister beetles, occasionally are involved in pollination. The larvae of some species are parasitic
in bee nests; others are parasitic on grasshopper egg masses. All adult Meloidae feed on pollen or on
both nectar and pollen. The larvae of
some species of Cleridae
are flower inhabiting. They are
mainly parasites in the nests of wasps and bees. The adults are predaceous, but they also feed on pollen. Most Melyridae are predaceous as
larvae and both predaceous and polleniferous as adults. One genus of Buprestidae,
Acmaeodera) (flat-headed borer) is polleniferous. The larvae bore into wood and the adults
feed on pollen. Many genera of Cerambycidae,
or long-horned beetles and round-headed borers, can be involved as
pollinators. The larvae bore into
wood but the adults feed on pollen.
Several genera of Scarabaeidae,
or white grubs, visit flowers. They
are primarily root-feeders as larvae, but they also feed on pollen as
adults. Elateridae,
or click beetles, are mostly root-feeders as larvae, but adults will feed on
nectar and pollen. In the Dermestidae,
the genus Anthrenus feed on decaying animal matter as larvae, but
adults may also utilize pollen (especially Anthrenus). There are also other small families of
Coleoptera, such as the Mordellidae,
Oedemeridae,
Lycidae
and Rhipiphoridae, whose members have
been observed to act as pollinators. Most groups of flowers do not
escape visits by beetles feeding on their petals as well as nectar and
pollen. Some blister beetles will
feed on legume petals in order to expose the pollen and nectar. Some very tiny flower-visiting beetles may
crawl into the narrowest corollas or tightest keels. Nevertheless, only a few groups of flowers
are visited regularly by a variety of beetles. Examples are flowers with abundant pollen, social flowers with
concealed nectar, flowers with exposed nectar and flowers with partially
concealed nectar. The adults of several large
families of Diptera feed
frequently on nectar or pollen or both, but the larvae are usually harmful to
plants. Examples are found in the Anthomyidae
(hovering house flies), Bombyliidae
(bee flies), Calliphoridae
(blow flies & bottle flies), Ceratopogonidae (biting midges)
Conopidae
(thick-headed flies), Cyrtidae
(small-headed flies), Empididae
(dance flies), Muscidae
(house flies), Sarcophagidae
(flesh flies), Stratiomyidae
(soldier flies), Syrphidae
(flower flies, syrphid flies, hover flies), Tabanidae
males (horse flies), Tachinidae
(tachinid flies), Tephritidae
(fruit flies). These families might
be considered in the following order of decreasing importance: Syrphidae, Muscidae, Calliphoridae,
Sarcophagidae, Bombyliidae, Conopidae, Tachinidae, Empididae, Stratiomyiidae,
Tabanidae, Tephritidae, Ceratopogonidae and Cyrtidae. However, this order may differ for any
one-plant species. A few of the more important pollinating Diptera are
discussed in the following. Syrphidae have larvae with a wide variety of habit. They occur under bark, manure and liquid
and are predatory on small insects such as aphids. The adults re almost all flower visitors. Most species feed on nectar and pollen or
only nectar. Nectar-feeding species
have a long, slender proboscis and generally visit the same group of flowers
as the long-tongued bees. Those
syrphids with short or moderate tongue length visit predominantly flowers of Groups I, Group II and Group III. Some also consume pollen on flowers of Group V. Bombyliidae have larvae that either feed on grasshopper egg
masses or those that feed on the larvae of wasps and wild bees. Adults of the latter group have a long,
slender proboscis and visit flowers of Group
III to Group VIII, but mostly Group III and Group IV. Although a few genera are intermediate,
most have very a short proboscis and visit primarily flowers of Group II. Muscidae have larvae with various habits. Some are internal parasites of other
insects, while some feed on plant roots, and a great many feed on decaying
animal and plant material. The adults
of most species visit flowers and eat pollen and nectar. Flowers of Group II are favored, but a few
others like onion in Group IV are
also visited. Species in other families of
Diptera will on rare occasions pollinate plants either directly or
accidentally. Adults of most Lepidoptera feed mainly on nectar
from flowers, while their larvae feed on herbage, some roots or stored food
products and wool and are therefore pestiferous. Their preferred flowers are in Groups IV to VII. Encounters with Hymenopterid flowers (Group
VI) often do not expose
the pollen and therefore do not result in pollination. The tongue lengths of Lepidoptera
vary from 1 to 250 mm. Those with
4-10 mm. Tongues are most often seen on flower Groups IV & V, while those
with longer tongues are most apt to be seen on Groups VI & VII. Butterflies tend to frequent
day-blooming flowers and moths visit constantly open or evening and
night-blooming flowers. The entire
suborder, Rhopalocera
and 5 families of Heterocera that are numerous or specially
adapted as pollinators are Arctiidae (tiger moths & wooly
bears), Geometridae
(loopers), Noctuidae
(nun moths, cut worms), Pyralidae
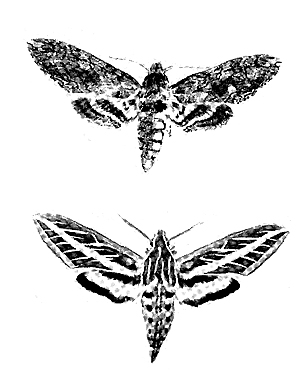
(snout moths), Rhopaloceridae (butterflies) and Sphingidae
(hawk moths & horn worms). Because investigations of visits
to flowers have been made primarily in daylight, the value of moths as
pollinators is probably underestimated.
Butterflies often spend a lot of time on the same flowers and they are
regularly less effective than bees in pollination. Haw moths that fly in the evening or at night are assiduous
flower visitors by darting rapidly from plant to plant. Their very long proboscis seems to be
especially suited for the most highly developed Lepidoptera flowers that have
musky odors, long and narrow corolla tubes or long spurs that contain
nectar. Butterflies tend to prefer
red flowers while moths prefer white flowers. Nun moths are similar to haw moths in rapid flight and long
tongues. They are usually more
abundant also. Many flowers are
sometimes referred to as haw moth flowers, and where the corolla tube exceeds
25 mm. the term is deserved. But,
hummingbirds and honeybirds contribute more effectively to the pollination of
such flowers in some areas. A small order, Thysanoptera
are tiny but individual species occur in large numbers. Adults and larvae feed either mostly on
honey and pollen or are predators of other thrips in flowers. It has been suggested that few indigenous
flowers in Europe escape from occasional or frequent visits by thrips. Even though individual thrips may only
convey pollen accidentally, their great abundance enhances their value for
pollination. Nevertheless, they are
generally thought to be ineffective in the pollination of many flower species
and consequently they are rarely credited with much influence. They rarely migrate from plant to plant so
that their role would be primarily self-pollination. Generalizations On
The Use of Non-Hymenoptera Pollinators
Among the Coleoptera, larvae of most species are destructive and not
advisable for propagation. One genus
of Cantharidae (Chauliognathus) are predators as larvae on aphids and
as adults they feed on nectar and pollen.
It is able to trip alfalfa and might be adaptable to mass production
in insectaries and mass release in field crops. The larvae of many genera of Diptera are destructive. Adults may
pose a health hazard and are thus unsuitable for purposeful deployment. Muscidae may be useful in confinement for
breeding work and small-scale increase of desirable plant stocks. There are may good pollinators among the Syrphidae,
however. They could be increased
rapidly and used as predaceous forms in insectaries. Although species may
resemble bees and wasps, they are non-biting. Semi-aquatic species could be
increased in field crops. The drone
fly, e.g., is an efficient fruit pollinator and might be propagated in
shallow tanks infused with organic material. Most larvae of Lepidoptera are also destructive and
thus the group is mostly unsuitable for deployment. There may be some exceptions, but any species considered would
need to be carefully studied for any possible destructive tendency. Vanessa cardin & V.
atalanta (L.) feed on thistles as larvae and might be considered for the
pollination of some ornamentals.
Sphinx moths are more destructive to weeds than crops (excluding
grapes) and could be deployed to pollinate ornamental plants. Vanilla is usually hand-pollinated, and
the search for a nondestructive Lepidoptera might be made. The possibilities for deploying
Lepidoptera as pollinators are probably greatest for agriculture in tropical
regions. |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}