|
File: <tachi1.ima.htm> [For educational purposes only] Terminology Glossary <Principal Natural
Enemy Groups > <Citations> |
{Please also see: http://www.tachinidae.org.uk}
Immature
Stages of Tachinidae
Clausen (1940) discussed the immature stages of Tachinidae as
follows: The Egg.‑‑There are
four general types of egg produced by the Tachinidae: 1.
Macrotype.‑‑The eggs of this form have a very
heavy and opaque dorsal chorion, with the ventral surface flat and the
chorion thin and more or less transparent.
They differ from other types principally in size, color, the stage of
development of the embryo at the time of deposition, and the manner in which
hatching is accomplished. The general
form is oblong, the anterior end being very nearly as wide as the posterior.
and in lateral view they are markedly arched, at times approaching the
hemispherical with the ventral side flat or at times slightly concave. A few species of Nemorilla and Gymnosoma
bear a distinct marginal flange at the juncture of the flat ventral surface
with the lateral areas. In size, the eggs of the different species range from
0.4 to 0.9 mm. in length, with the width varying from one‑third to two‑thirds
the length. The color is usually
creamy‑ or glossy‑white at the time of deposition, later changing
to a gray or light brown. The
micropylar area is usually dorsal and somewhat pigmented, though in Gymnosoma it is terminal or
slightly ventral. In a few species,
the surface of the chorion bears rather minute reticulate markings. Townsend
described the unusual egg of Phasiopsis
floridana Tns. as
asymmetrically long‑ovate, the right side straight on the middle half
and the left side evenly curved, and brown in color. It differs from those of other species,
also, in revealing this coloration even before deposition. The macrotype eggs are divided, on the basis of the manner of
hatching, into two distinct groups termed "dehiscent" and
"indehiscent" by Pantel.
The latter form has no special provision for hatching, which is
accomplished by cutting through the thin ventral shell directly into the host
body. The eggs of Centeter, Meigenia, and Trichopoda
are representative of this group. The
dehiscent eggs have a distinct seam, or line of fracture, across the anterior
margin, which curves backward and somewhat dorsad; at hatching, this seam is
broken, and the lid is forced upward to permit the larva to emerge into the
open. Ptychomyia, Tricholyga,
and Winthemia (Fig. 209F)
deposit eggs of this form. The macrotype eggs carry a quantity of
mucilaginous material at the time of deposition, which fastens them securely
to the body of the host. In some
species, this material may be observed in regular loops or folds on the
venter of the uterine egg. Please
CLICK on picture to view details:
2.
Microtype.--This type of egg has many characters in common with the
macrotype; as the name indicates, it is of minute size in relation to the body
of the parent female. One of the
largest recorded is that of Chaetogaedia
monticola, which measures
0.44 by 0.25 mm. Thompson has given
the measurements of a large number of eggs of this type, the great majority
of which range from 0.02 to 0.2 mm. in length. Those of Zenillia
pullata and Pseudogonia cinarescens Rond. are
exceptionally minute, measuring 0.027 by 0.02 mm. and 0.08 by 0.05O mm.,
respectively. The majority are
approximately two‑thirds as wide as long, ranging to almost circular in
Phryno vetula Meig.
Townsend calculated the volume of a considerable number of eggs of
this family and reported that the large macrotype egg of Gymnosoma, which is 0.9 mm. long, is approximately two
thousand times the volume of the microtype egg of Z. pullata. In general, microtype eggs are only about
one‑fiftieth as large by volume as macrotype eggs deposited by females
of the same size. Please
CLICK on picture to
view details:
The general form of the egg is ovate, with the anterior end
narrower. In side view, the outline is
somewhat semicircular, though often somewhat flattened; but in Racodineura antiqua (Fig. 207C) (Thompson,
1920b, 1928) the dorsum is in the form of an asymmetrical cone, with the
summit bearing an irregular group of strongly sclerotized elevations, which
are surrounded by a number of small button‑like structures, doubtfully
considered to be respiratory papillae (Clausen 1940). With reference to Otomasicera patella
Tns., the egg is described as limpet‑like, with irregular concentric
peripheral thickenings. Those of many
genera of Exoristinae and of Epidexia,
also, are very elongated, subelliptical, and pointed at both ends; they have
submembranous chorion extensions and the dorsum is black, coarsely punctate,
with reticulate markings and a median longitudinal suture. The surface markings of the eggs of other species range from a
very fine punctuation to a deep pitting or reticulate marking. The heavy
chorion of the dorsum ranges in color from gray to brown and to the more
frequent shining black, though Townsend listed several genera in which they
are yellow or even white. In Pexopsis aprica Meig., there is a densely pigmented hemispherical
process of unknown significance, at the anterior end; the egg of Sisyropa sp., which has the
reticulate surface, bears also an irregular, light‑colored fringe
around the margin, which is pierced with microscopic holes. On the ventral side, the chorion of the
microtype egg is thin and transparent, though in Gonia capitata
(Fig. 207B) the thickening and pigmentation are only reduced ventrally and
are entirely lacking only on the median line. The micropyle is clearly
recognizable in the eggs of many species and occurs at the narrower,
anterior end. The embryo is enveloped
by a tough vitelline membrane. 3.
Membranous.--This type of egg is distinguished in general by
its elongated form and thin transparent chorion. In size it is minute to large, ranging from 0.2 mm., about half
the length of the largest microtype egg, to equal to the macrotype eggs from
equal sized females, and the length may be from three to six times the
width. The ventral surface is not
flattened and therefore does not permit of close attachment, to the host or
to foliage. Its form may range from
long cylindrical, with both ends evenly rounded, to markedly arched or bowed,
with the ventral line straight or concave and the anterior end appreciably
narrowed. The micropyle is terminal
and may be simple, of rosette form, or with various short processes, which
are particularly noticeable on the ovarian eggs. The surface reticulations of
the chorion, termed the "pneumatic apparatus" by Pantel, are often
quite conspicuous, the lines being brownish in color. These reticulations may be complete, as in
Bonnetia comta (Fig. 207D), limited to
the middle portion of the egg, or of uneven distribution as in Peleteria prompta Meig. These thin‑shelled eggs hatch in the uterus of the female
or almost immediately after deposition.
The few species that deposit unincubated eggs of this type inject them
into the body of the host. 4.
Pedicellate.--The egg of Carcelia
(Fig. 207E) is placed in this group.
It has a membranous chorion and bears a slender cylindrical pedicel
about 1/5th the length of the body, at the posterior end. This stalk is expanded at its distal end
into an adhesive process, by means of which it is fastened to a hair or to
the integument of the caterpillar body.
First‑instar Larvae.--There are three general forms of
first‑instar larvae follows: 1.
Tachiniform.--The larvae included in this group are those
which may bc considered as normal for the family, and they have no marked
modifications in form or structure to adapt them to a specialized mode of
life. as number of larvae of this type have been described in detail by
Thompson (1926). They include all
those hatching from macrotype and pedicellate eggs, those which are injected
into the body of the host, and finally, those from the membranous type of egg
which, as fully incubated eggs or newly hatched larvae, are deposited upon
tho host body or in its immediate vicinity. The latter are presumed to reach
the host very quickly and consequently require no modification of the
integument to protect them from desiccation and injury. The body is usually robust, with the
cuticle colorless and transparent, and usually bears bands of rather minute
spines on each segment, particularly at the margins, these bands being most
frequently complete on the thoracic segments and predominantly ventral on the
abdomen. A few species, such as Centeter cinerea (Fig. 208A) appear to lack the spine bands
entirely. A definite adaptation for
attachment to internal tissues or organs of the host is found in Anetia, Compsi8lura, Paradexodes,
and some other genera. This consists
of a set of three heavily sclerotized spiracular hooks at the caudal end of
the body, each of the two dorsal ones being situated immediately above a
spiracle and the third on the median line somewhat below them. The dorsal hook3 are simple and sharply
curved, with the points directed cephalad, whereas the median one is double‑
or triple‑pointed and directed ventrad. These hooks are integumentary in origin and are not a part of
the spiracular structure itself, thus differing from the spiracular hooks in
later instars of certain other species.
In this group is also included the vesiculate larva of Plagia trepida, which, if the adaptation were more highly
developed and of more frequent occurrence, might constitute a separate type,
as in the Hymenoptera. 2.
Microtype larva.--These larvae, which are derived from microtype
eggs that hatch within the digestive tract of the host, are of very small
size and lack the various adaptations associated with a period of free life
or with the necessity of penetrating n heavily chitinized host
integument. The buccopharyngeal
armature usually shows a reduction and simplification. Thompson (l924) has given detailed descriptions
of the larvae of a large number of species, with keys for distinguishing them. The skin is thin and colorless and without
any evidence of specialization except for the transverse rows of minute
spines at both margins of the body segments, those on #he abdomen usually
being present only on the venter (Fig. 208E). In Gonia capitata and various other
species, the first segment bears dorsally at its anterior margin a group of
I0-12 strong hooks, which are heavily pigmented. In several species of Gonia
described by Tothill, there are transverse rows of four hooks ventrally
between the posterior segments, and the outer ones are connected by an
internal chitinous rod. The larvae of
Racodineura antiqua, Brachychaeta spinigera
Rond. and Pales pavida Meig. lack the caudal
spiracles, whereas in Phryno
vetula they are present but
very small and probably not functional.
The spiracles arc lacking also in Exorista
fimbriata Meig., and the
felt chamber is short, filiform, and almost invisible. The frequent absence of an open tracheal
system in larvae of this type is correlated with their mode of life in this
stage, during which they are usually embedded in some host organ. Except for its size and its association
with the microtype egg, the microtype larva has few characters to distinguish
it from the tachiniform larva. 3.
Planidium larva.--The essential adaptive characters of the
tachinid planidium are for the purpose of protection from injury and
desiccation during a more or less protracted period of free life before the
host is reached. They consist of a
cuticular armature of closely set polygonal sclerotized plates, or imbricated
scales, covering the dorsum and pleural areas, and in some species a large
portion of the venter as well, of all body segments except the last, forming
a carapace. Though most planidia have
the armature in the form of scales or plates, yet in several species it is in
the form of nodules or minute rounded protuberances surmounted by setae. Rows of spines occur ventrally upon the
various segments. In Fortisia foeda, each body segment except the last bears many small
oval plates, each of which terminates in a strongly sclerotized tooth
(Thompson, I915c). The extreme development of this type of larva in the
family is found in Gymnocheta
alcedo Lw. described by
Thompson (1923d), in which each segment bears a single dorsal plate and a
pair of pleural plates, and in species of Ormia,
Ormiophasia, and Euphasiopteryx described by
Townsend. The segments of the
planidia of the latter genera are telescopic, and the plates occur in three
series, the dorsal one being very wide and the others situated dorsolaterally
and ventrolaterally. The venter is
unsclerotized. Ormiophasia still further approaches the planidium type in
that it possesses caudal cerci, a character found in the larvae of many
Dexiinae. The planidia are derived from eggs hatching in the uterus of the
female or from membranous eggs deposited on the food plants of the host or in
their general vicinity. They are
variously colored, owing to the pigmentation of the plates, and may be grayish‑white,
brown, bluish‑green, or black.
As growth takes place, the plates become considerably separated,
revealing the white body color. There is also an appreciable stretching of
the thin intersegmental membranes, resulting in a distinctly banded appearance. This is noticeable even in the freshly
deposited larvae of A analis. The species that retain the eggshell as an
anchor to the substratum, and enveloping the caudal end of the body, have
forwardly directed spines on the last one or two segments. Typical of this group are Archytas analis, Bonnetia
comta (Fig. 208C,D), and Ernestia ampelus. The first‑instar
larvae of Ophirionopsis
and Ophirion are markedly
different from other representatives of this group. They are described by Townsend (193fi) as being somewhat
caterpillar‑like and very active; the former is stated to have eight
pairs of "pseudolegs," which are half as long as the thickness of
the body, and it has, in addition, three anal pseudopods. In Ophirion,
the pseudolegs occur on the 4th to the 10th and on the 12th and 13th
segments. The occurrence of light‑colored planidia is exceptional and
a number described as such are believed to represent larvae deposited
prematurely or dissected from the uterus and mistakenly believed to be
mature. All larvae of this type have
open caudal spiracles. The three first‑instar larval forms described above are
separated on the basis of adaptive characters, which are of independent
development in widely separated genera and higher groups. The morphological characters of value in
classification require too detailed treatment to be adequately summarized
here, but in general they relate to various details of the head structure,
the buccopharyngeal apparatus, the arrangement and form of the integumentary
spines, the sensory organs, the caudal spiracles, etc. Principal studies upon the classification
of these larvae have been by Thompson (1922; 1923b,c; 1924), who concluded
that they present adequate characters for specific determination, though the
groups set up on this basis are not always in accord with those based upon
the taxonomy of the adults. He
pointed out further that it is often difficult or impossible to distinguish
between larvae of species which are quite distinct in the adult stage and, conversely,
that it is at times possible to distinguish definitely between larvae of
forms which, in the adult stage, are apparently identical morphologically
(varieties of Lydella stabulans Meig.). In practically all first‑instar larvae, the number of abdominal
segments recorded has been eight.
There is a considerable range of variation in certain of the
morphological characters that are not associated with a particular type of
larva but that are used in the classification of the smaller groups. Some of
these are discussed briefly herewith. The buccopharyngeal apparatus consists, with very few exceptions,
of a simple, unjointed structure, of which the three principal parts are the
median tooth and the intermediate and basal regions, the latter of which may
be only lightly sclerotized. The
relative lengths of these three parts vary greatly. The outer, or dorsal, margin of the tooth may be smooth or bear
a number of teeth. The lower wings of
the basal region are deflected somewhat ventrally, whereas the upper ones are
usually considerably arched. There are frequently small lateral plates at
the sides of the median tooth, and the small salivary-gland plate, often
delicate and inconspicuous, lies beneath the intermediate region. The anterior lateral plates at each side
of the median tooth can frequently be recognized, though they may be very
lightly sclerotized. A distinctive form of the buccopharyngeal structure is
found in Bigonichaeta setipennis in which the
intermediate region is in the form of a straight elongated rod, quadrangular
in cross section eight times longer than wide. The basal region is short and feebly developed. The sensory organs of the head are usually not greatly developed
and are most conspicuous in Bigonichaeta. In addition to the usual maxillary
sensoria, there is a pair of prominent clavate sensoria dorsally just in
front of the antennae. The antennae
are themselves remarkably developed, being cylindrical and about six times as
long as wide, tapering abruptly to a point, and terminating in a hair‑like
distal portion about 4X as long as the basal part. In addition to the sensory setae of the body segments, there
are circular sensory organs on the venter of the various segments. In some
species, such as Argentoepalpus
signifer Wlk. and others
described and figured by Thompson, there are rod‑ or club‑shaped
sensoria an the thoracic and last abdominal segments. In many species, the
antennae are small and somewhat conical in form, without a terminal hair. The tracheal system of the first‑instar larva, which is
present in the very great majority of species, consists of two main
longitudinal trunks, with relatively few branches; they are connected by a
posterior commissure. The posterior
spiracles usually consist of two papillae, each spiracle consequently being
kidney‑shaped in outline, though a few species have simple circular
spiracles with only a single opening.
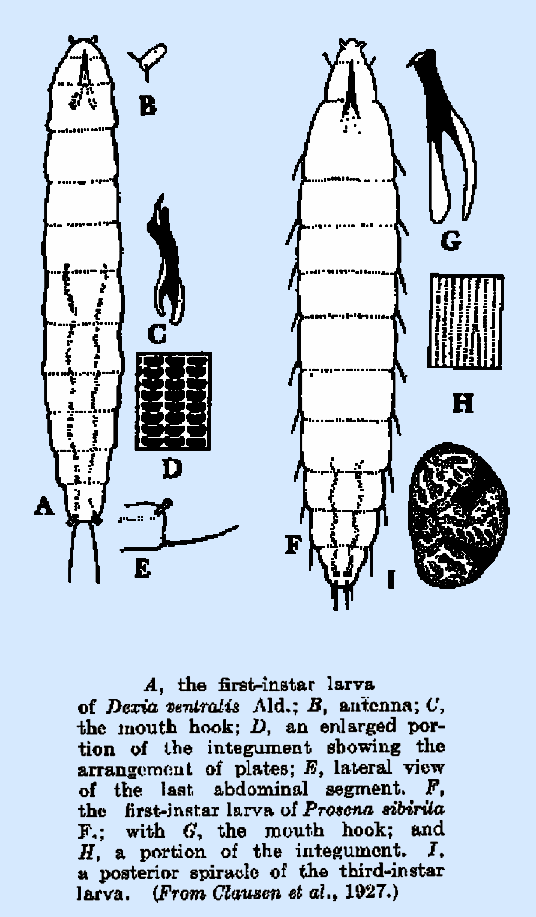
In Dexia ventralis (Fig. 209E) and Theresia claripalpis, the spiracles are borne upon short
cylindrical stalks; in Billaea
pectinata Meig. (Tolg,
1910), the stalks equal one body segment in length, and they also bear three
long setae at the distal end. The
atrium, spiracular, or "felt" chamber may be only as long as wide (Leschenaultia exul) or range to 15 to 20
times the width (Bigonichaeta
setipennis). It has been mentioned that spiracles are
lacking in a number of species of microtype larvae, and they are likewise
missing in Actia diffidens and Sturmia pelmatoprocta B. & B.
Anterior spiracles are quite uniformly absent in the first instar,
though Landis states that they are present in Paradexodes epilachnae. Second‑instar
Larvae.‑‑The larvae of this instar show a much greater
uniformity than do those of the preceding instar, as is expected in view of
the occurrence of all species in the same environment, that is, within the
body of the host. The factors of
locomotion, desiccation, penetration, and mechanical injury are all absent;
and consequently adaptations to meet these conditions are lacking. In all species, the integument is thin and
transparent, the most conspicuous change in this respect being among the
planidium‑type larvae, which discard the heavy integumentary armature
of plates, scales, etc., at the first molt. Generally, the cuticular armature of this instar consists of
bands of setae, usually rather delicate, about the thoracic segments, on the
venter of the abdominal segments, and in the form of a large patch on the
last segment, often accompanied by a rather heavy band on the penultimate
segment. These setae are often
arranged in irregular and broken rows.
In some species, the spines are much more numerous than in the
preceding instar; in others, they are much less conspicuous. Several species having heavily spined
planidium‑type larvae are, in this instar, virtually devoid of
spines. In several species, rather
conspicuous departures from the normal spine arrangement are known. The larva of Gonia capitata
has patches of minute black spines on the dorsum of the first thoracic
segment and on the venter of the second, which are so dense as to give the
appearance of black plates. The spine
patches on the venter of the last abdominal segment reach a maximum
development in Anetia hyphantriae and are said to
invaginate to serve as false feet, though the locomotory requirements of this
instar are quite limited. The spines
of the last two segments of the great majority of species are usually
directed cephalad, and they serve to hold the larva more firmly in position
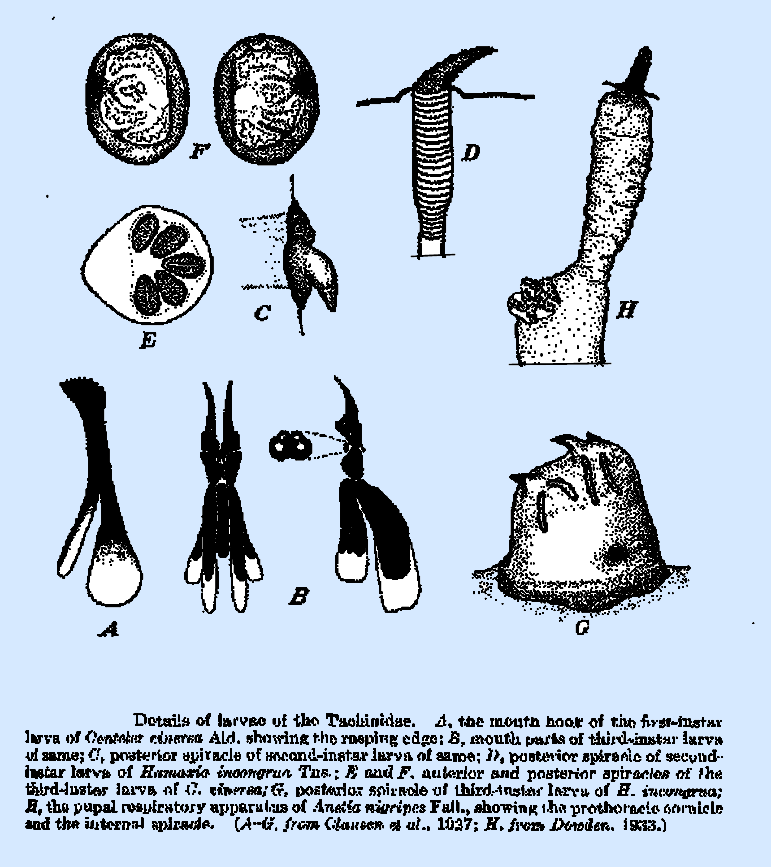
in the respiratory funnel. The larva of Centeter
cinerea is distinguished
from others of this instar by the possession of a pair of conspicuous anal
lobes which may be homologous with those of the same instar of certain
Conopidae. The buccopharyngeal apparatus is more robust and highly developed
in the second than in the first instar, and there is, of course, much
variation in the form, relative size, and sclerotization of the different
parts. In all species, the paired
mandibular hooks are present, in contrast to the single median tooth of the
first instar. A few species, such as Zenillia
libatrix, Bigonichaeta setipennis, and G. capitata, have no articulations, the entire structure
apparently being fused into a single piece.
The majority, however, have one articulation, which is usually between
the anterior and intermediate regions, though in some it is between the
intermediate and basal regions. Archytas analis, Bonnetia
comta, and Siphona geniculata have two articulations, separating the three
principal parts of the structure. The respiratory system of the second‑instar larva is also
much more highly developed than in the first instar, because of the greater
need for oxygen. The two longitudinal
trunks are heavier, with a considerable number of branches, and both
posterior and anterior commissures are present. In about half the species that have been studied, the anterior
spiracles are said to be lacking, but in many instances they have probably
been overlooked; for they are often very minute and situated intersegmentally
in the pleural area between the first and second thoracic segments, and, in
this position, any shrinkage or contraction of the body would tend to obscure
them. Each anterior spiracle usually
has two or three papillae, the number being increased to 3-5 in Bigonichaeta setipennis, 5-6 in S. geniculata, and 6-9 in Lydella
stabulans. In Ernestia
ampelus, however, they
appear as simple circular openings, and those of Billaea pectinata
are mere slits in the integument, and are recognizable only in prepared
sections. The posterior spiracles of the majority of species are widely
spaced and have two papillae, lobes, or spiracular slits, though Actia diffidens has only one and Leschenaultia exul
and Siphona geniculata have three. An exceptional modification is present in
the posterior spiracles of Centeter
spp. (Fig. 210C) in the form of a large, ventrally directed, conical process
immediately beneath the openings, which is believed to aid in perforating and
maintaining connection with an air sac of the host. This process in Hamaxia
incongrua (Fig. 210D) is
more sharply pointed and directed dorsad.
In Anetia and related
forms, the stigmatic hooks, described for the first instar, persist in
somewhat modified form. That they are functionless is improbable, in view of
the development of a respiratory funnel during this stage. The spiracular chamber in most species is
very short, often being broader than long.
The only instance of complete lack of spiracles in the second‑instar
larva is that of Fortisia foeda cited by Thompson, though
the internal tracheal system is present and the trunks are filled with air. Third‑instar
Larvae.‑‑The mature larva is usually somewhat crescentic in
lateral view, with the venter concave and the abdominal region broadest. In some species, as Chaetogaedia analis
v.d.W., the caudal segment is approximately the same width as those preceding
it, whereas in others there is an appreciable tapering caudad. The segmentation is distinct, though
frequently obscured by segmental folds.
The larvae of Zenillia
roseanae and Actia diffidens have distinct median pseudopodia ventrally
between the abdominal segments. The
anal opening occurs at a variable distance beneath the posterior spiracles,
often near the anterior ventral margin of the last segment. The integumentary armature, consisting of
spines and hairs, may be somewhat more extensive than on the preceding
instar. The larvae of Bonnetia comta, Bonichaeta
setipennis, Racodineura antiqua, Dexia ventralis,
and Prosena sibirita are almost bare, whereas
that of Sturmia inconspicua is almost
completely covered with setae. More
frequently, however, the thoracic segments and the last one or two abdominal
segments bear complete bands of setae, whereas on the intervening segments
they are largely ventral. The setae
of the anterior segments are directed caudad and on the posterior segments
usually cephalad. In Centeter cinerea, the central area of the last segment is covered
with a patch of heavy black spines.
The so‑called spiracular hooks of Zenillia spp. persist in the form of clusters of three or
four black spines, the bases of which are fused. These are apparently homologous with similar groups found in
the same position on certain conopid larvae.
The sensory organs are often reduced in size and number as compared
with those of the preceding instars. The four pairs of finger‑like
organs on the last segment persist in Zenillia
libatrix. The buccopharyngeal apparatus is more robust and highly developed
than in the preceding instars. The
majority of species now have distinct articulations separating the three
principal regions. A considerable
number of species, however, have only one articulation, which in some cases
occurs between the anterior and intermediate regions and in others between
the latter and the basal region. Bigonichaeta is distinctive in
having the entire structure in one solid piece. It is thus seen that the third‑instar larvae of the
family normally have two articulations, but frequently only one and rarely
none; the second‑instar larvae most frequently have one, but
occasionally two or none; and the first‑instar larvae usually have
none, but very infrequently one or two.
In no instance is a smaller number of articulations present than in
the preceding instar of the same species.
B. setipennis and a species given
by Nielsen as Ernestia connivens Zett., but later
stated to be Plagia trepida instead, have no
articulations in any instar; Zenillia
spp. have none in the first and second and one in the third instar; Lydella
stabulans has none in the
first and one in the following two instars; and Leschenaultia has none in the first two and two in the
third instar. Several species have
none in the first and two in the last two instars. The greater number of species have none in the first, one in
the second, and two in the third instar. The anterior region is quite generally in the form of paired
mandibles which are distinctly hooked.
The hypopharyngeal and epipharyngeal sclerites are situated
immediately beneath the juncture of the anterior and intermediate
regions. Each of these bears clear
areas which are believed to represent sensory organs. Those of the
epipharyngeal sclerite particularly are variable in number and form and are
considered to be of value in making specific determinations. The respiratory system reaches its greatest development in the
third‑instar larva, as is to be expected from its greater size and the
probable complete cessation of cutaneous respiration. It consists of a pair of heavy
longitudinal trunks, anterior and posterior commissures, and anterior
(occasionally absent) and posterior spiracles. In Bonnetia comta, each trunk is reported
to have a diameter equal to one‑fourth that of the body. According to Rennie & Sutherland
(1920), Siphona geniculata lacks the anterior
commissure. The anterior spiracles are situated dorsolaterally at the
posterior margin of the prothorax and may be in the form of a single circular
opening as in Winthemia quadripustulata; a circular
plate bearing radiating slits as in Centeter
(Fig. 210E), Hamaxia, and Bonnetia; or, more commonly, a
conical process bearing a number of papillae. These papillae may range in number from two to three in Zenillia spp to 30-35 in Billaea pectinata.
Occasionally, as in Siphona
and Racodineura, the
spiracle is fan‑shaped, with the papillae in a rows at the outer
margin. In Archytas analis
and G. capitata, the spiracular chamber itself is bifurcate, with
an opening at the end of each short branch.
In the former, the openings are elongate and curved. The spiracular chamber is usually two or
three times longer than wide. The
anterior spiracles are not believed recognizable if present in Sturmia inconspicua (Webber 1932) and are lacking in Leschenaultia exul, though the stubs of the
tracheal branches are present. The posterior spiracles are usually large, somewhat circular in
outline, with the inner margins more or less flattened, occasionally almost
straight, and encircled by highly sclerotized, usually black,
peritremes. In some instances, these
spiracles are semicircular or even triangular in outline. The peritremes may be shallow, or they may
exceed the width of the spiracle, forming conical or subcylindrical
bases. Occasionally, as in Siphona and Ernestia rudis, they are incomplete, being broken on the inner
margin. In the great majority of
species, the spiracles are situated above the transverse axis and
occasionally are distinctly dorsal.
Very seldom are they separated by a distance greater than the width of
one spiracle, and frequently they are almost contiguous. The spiracular slits are usually straight
or slightly curved and radiate outward from the spiracular scar, which itself
is situated at or below the median transverse line and somewhat toward the
inner margin. The number of these
radiating slits is variable, most frequently being 3 or 4 but ranging up to 6
in Eubiomyia calosomae and about 10,
variable and often branched, in Bigonichaeta.
In Racodineura, there are 30
short slits arranged side by side in the peripheral part of the respiratory
area. Several species, such as Gonia
capitata, show the smaller
number of slits following the periphery rather than radiating from the
vicinity of the scar. An occasional
species has exceedingly long, serpentine slits, which at times are
extensively branched. The three
elongated slits of G. capitata and Sturmia inconspicua occur at the crests of pronounced ridges. An unusual modification in form of the posterior spiracle is
found in H. incongrua in which the face of
the spiracle (Fig. 210G) is rounded and highly sclerotized, forming, with the
peritreme, a broadly rounded cone surmounted at one side by three hook‑like
spines, which are directed laterad.
In Carcelia gnava, the respiratory area is
in three parts, with the openings small, irregular in form, without apparent
order, and numbering about 30. The spiracle of Steiniella callida
Meig. has 80-100 minute openings (Nielsen, 1909), and that of Oedematocera dampfi Ald. is rosette‑shaped
with the pores arranged in rows radiating from the center (Greene,
1927). The spiracles of Fortisia foeda are very large and bulbous and surmount short, broad
stalks. The openings are small,
irregularly placed, and very numerous.
A remarkable form is figured by Lloyd for Ginglymyia acirostris,
in which the large spiracular stalks, which are oval in cross section, arise
from a common base, definitely dorsal in position, and each one terminates in
an elongate‑oval spiracle with a single elongated, curved opening at
the center (Fig. 205D). At the outer
margin of the spiracle are 30 leaf‑like processes, of unknown
function. These spiracles represent a
wide departure from the normal tachinid form. In a number of species, several minute openings have been noted
on the surface of the spiracle, which are stated to be those of the
perispiracular glands. The spiracular
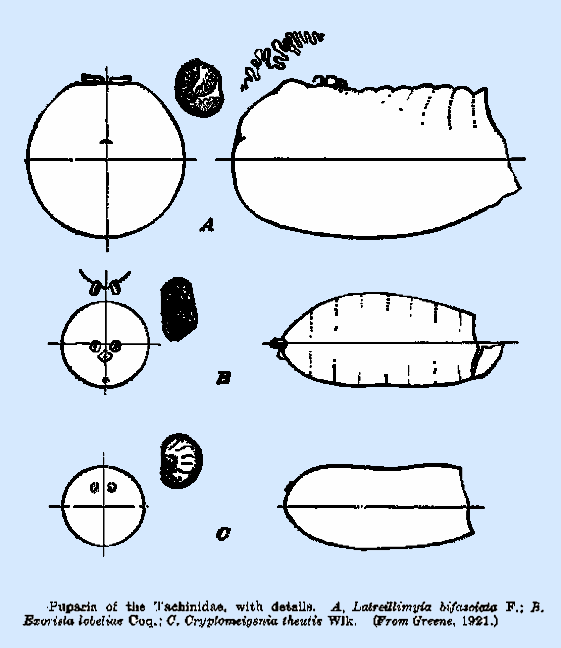
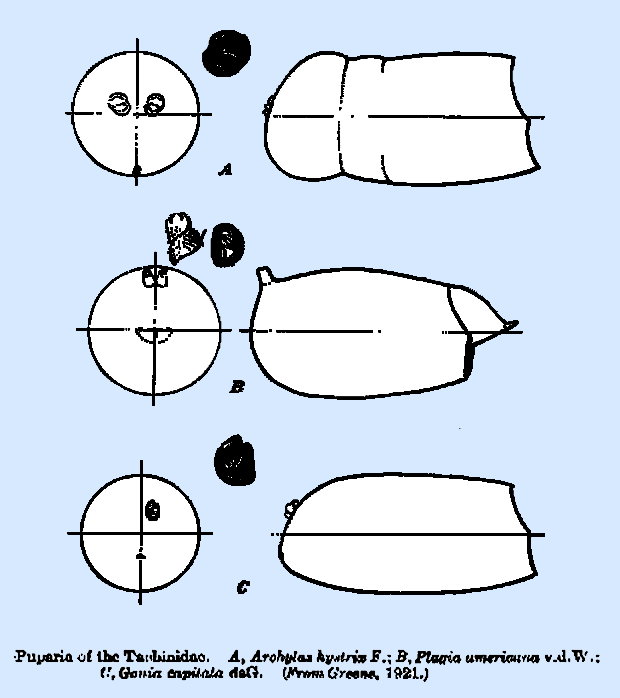
chamber is relatively short in most species, seldom being longer than wide. Puparia.‑‑There
is an exceedingly wide range of variation in form and general characters
among the puparia of the Tachinidae. Those of a considerable number of
species have been described in connection with biological studies of
particular species, but the most detailed account yet available is that by
Greene (1922), in which the puparia of 100 species, of muscoid flies, a large
portion of which are of this family, are described and figured. The author concluded that the puparial
characters are adequate for specific determination. The general form of the puparium is subelliptical and slightly
widest in the midabdominal area, with both ends smoothly rounded. In a few
species, such as Hamaxia incongrua, Cryptomeigenia aurifacies
Walt., and Viviania georgiae B. & B., the
abdominal region is much larger than the anterior, though occasionally it is
somewhat narrower. The posterior end
is at times markedly truncate, as in Sturmia
cubaecola and Chaetogaedia analis v.d.W., while in others
the caudal segments may he progressively narrower, giving a distinctly
pointed appearance. Usually, the
longitudinal axis is straight, though in Eutrixa
exile Coq. there is a marked
upward curve of the anterior region.
The segmentation is usually indistinct and is indicated by faint lines
or by a variation in the pubescence; yet in Exorista confinis
Fall. the intersegmental constrictions are very distinct on the entire
periphery, and in several species they are pronounced on the dorsum. In Pyraustomyia
penitalis Coq. and others,
the last abdominal segment is much reduced, forming a tubercle surmounted by
the spiracles. The anal opening is
represented by a distinct groove at a varying distance below the spiracles,
frequently occurring almost at the anterior ventral margin of the last
segment. The color of the puparia of any given species is quite variable
and usually deepens appreciably with age.
In general, it ranges from the darker shades of red and brown to black
and may be dull or have a silken luster.
The puparia of a considerable number of species are yellowish‑red,
and that of Alophora pulverea Coq.is pale‑yellow. The outersurface of the puparium bears the
armature of the third‑instar larva and may consequently be hairy, bare
and smooth, or roughened by striations or rugosities. The anterior and posterior spiracles are, of course, those of the
third‑instar larva; and although they are still recognizable as such,
yet they show appreciable changes. The
posterior ones are generally situated slightly above the transverse axis of
the body; but they are definitely dorsal in Latreillimyia bifasciarta
(Fig. 211A), though they are below the axis in a considerable number of
species and conspicuously so in Tachinophyta
floridensis Tns. They may be only slightly raised above the
surface of the puparium or borne upon pronounced tubercles, as in Anachaetopsis tortricis Coq. The spiracular
slits correspond in form to those of the mature larva. The outstanding variation in spiracle form
and position is found in Ginglymyia
acrirostris, which bears them
on large and heavy individual stalks markedly dorsad in position. The puparium of Thrixion halidayanum
is distinguished by the occurrence of the
spiracles upon a common stalk, which is large and cylindrical and
projects from the rounded posterior end. The external prothoracic cornicles are lacking in the majority of
species, but in those which bear them they appear as conical or
subcylindrical projections through the puparial wall dorsolaterally near the
posterior margin of the fourth segment.
The papillae usually number 6-20, though in Leschenaultia exul
they exceed 100, distributed irregularly over the distal half of the
cornicle. In Actia diffidens
there is only a single terminal opening.
In Siphona geniculata, the perforations
through the puparial wall may be detected, though the cornicles do not
protrude. The internal prothoracic spiracles of the pupa (Fig. 210H),
situated at the base of the prothoracic cornicles and beneath the puparial
wall, are present in all species.
They are circular in outline and nearly flat and usually bear the
minute papillae in double rows radiating from the center. These rows are irregular and sometimes
branching and number 5-6 in Zenillia
libatrix. The total number of papillae in the great
majority of species is 100-200.to 200. There are two lines of cleavage which separate the two halves of
the puparial cap from each other and from the remainder of the puparial wall,
and both these halves are broken away at the time of emergence of the adult
fly. The horizontal line of cleavage,
separating the two halves, extends across the front and posteriorly at each
side to a point in the anterior portion of the fourth segment. The vertical line of cleavage passes
completely around the puparium and through these points and is in front of
the prothoracic cornicles of the pupa, if they are present. References: Please refer to <biology.ref.htm> Additional
references may be found at: MELVYL Library] |