|
File: <strep1.ima.htm> [For educational purposes only] Terminology Glossary <Principal Natural
Enemy Groups > <Citations> |
Immature Stages of Strepsiptera
Clausen (1940) discussed the immature stages of Strepsiptera as
follows: The Egg.‑‑The fully
developed eggs in the body cavity of the female are minute and almost
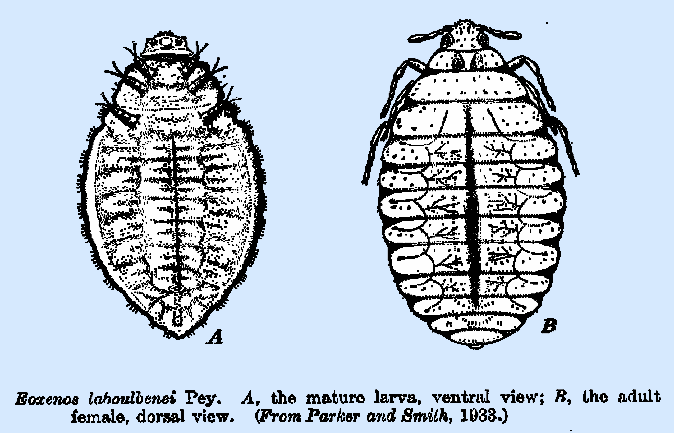
spherical in form and usually have the chorion thin and membranous. That of Eoxenos laboulbenei
is 0.07 mm. in diameter and, in its early stages, is seen to have a band of
irregular hexagonal cells extending around it. These are not surface markings, however, but represent the
developing embryo. In some species, at least, there is an increase in size
during incubation, and just prior to hatching the larva lies in a curled
position, with the posterior portion of the abdomen sharply bent ventrally
beneath the body. The
Larval Instars.‑‑The first‑instar larva of the
Strepsiptera is generally referred to as a triungulinid because of its marked
resemblance to the larvae of the Rhipiphoridae, which were originally
designated by that term. It is,
however, essentially a planidium, and various authors have called attention
to the similarity of its adaptive characters and habits to those of the
planidia of the Eucharidae, Perilampidae, Cyrtidae, etc. Pierce and other authors have described the triungulinids of a
considerable number of species, and the characters available appear to be
sufficient to permit determinations of the instar to the family. Further study may reveal a basis for
determination to lower groups. Please
CLICK on picture to view
details:
In general, the triungulinids are fusiform in outline with the
head and body segments heavily sclerotized, ranging in color from amber to
shining black, the segmental bands terminating in pleural plates
lateroventrally, the abdomen bearing one or more pairs of cerci or stylets on
the terminal segments and a caudal sucker. After feeding has taken place, the
bands become widely separated because of the stretching of the thin
intersegmental membranes and reveal the white color of the body contents. In size, the triungulinids are quite
small, seldom exceeding 0.3 mm. in length, and many are only half that
length. This is understandable in
view of the large numbers produced by each female. Please CLICK on picture to view
details:
The head in most species is somewhat semicircular in outline,
often slightly longer than wide, and occasionally is almost
quadrangular. The eyes are usually large
and situated slightly beneath the lateral margin. They consist of a group of three to five lenses, variously
arranged, which are underlain by masses of pigment. A few species apparently lack the lenses entirely. The antennae may be entirely absent, as in
Eoxenos laboulbenei, or occasionally disk‑like or slender
and two‑ or three‑jointed, as in Callipharixenos muiri
Pierce and Stichotrema dallatorreanum Hof. In many species, the mandibles are exceedingly minute and spine‑like
or entirely lacking, but in the last two species named they are rather large,
curved, and directed backward. E. laboulbenei has the axillae very long and spine‑like. The thoracic segments are longer than the abdominal segments
following and are strongly arched.
There is considerable variation in the 4‑jointed legs of the
various species, particularly in the tarsus.
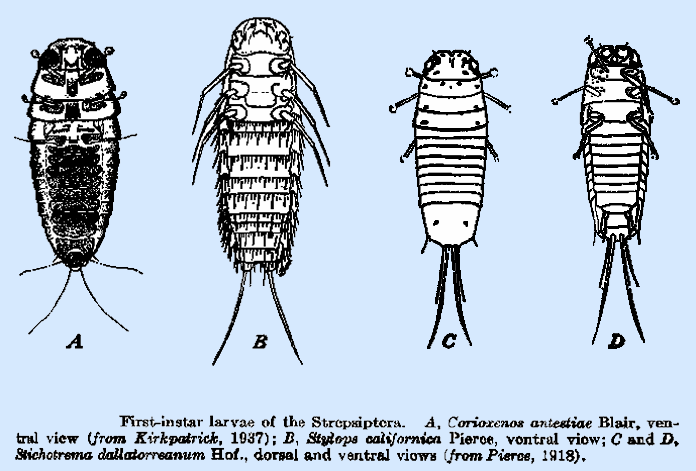
In Stylops californica Pierce (Fig. 218B),
the tarsi are spine‑like and equal in length to the tibiae, whereas in C. muiri they are only half as long as the tibiae and
terminate in three filaments or claws.
In the majority of species, however, the tarsi of the first two pairs
of legs are in the form of large disk‑like pulvilli, and those of the
hind legs are slender and tapering.
All three pairs of legs of Stichotrema
dallatorreanum (Fig. 218C,D)
are thought to have the pulvilli.
The unguliform tarsus of the hind leg of Xenos bohlsi
Hoff. is replaced by a ladle‑like pad.
The tarsi of all the legs of Stylops
swenki are described as
being acuminate in form and nearly half the length of the tibiae. The abdomen consists of 10 segments of gradually decreasing
width, though Callipharixenos
is thought to have only 7 segments.
The first eight segments are short and of approximately equal length,
whereas the terminal two are often considerably modified. In Stichotrema
dallatorreanum, the ninth
segment equals the preceding five in length, and the tenth is small and
situated beneath the ninth. The last
segment is frequently emarginate and bears two pronounced terminal tubercles
from which the stylets arise. Only a
single pair of caudal stylets is found in the majority of species, these
being one‑half to two‑thirds the length of the body, though two
pairs are mentioned for certain species of Xenos, Elenchoides,
Corioxenos, Belonogastechthrus, and Mecynocera, and three pairs for
Neocholax jacobsoni Meig. If two pairs are present, the anterior
pair is shorter and is usually borne on the ninth segment. The three pairs on Neocholax are all borne on the penultimate segment, and in
E. laboulbenei the two pairs are both on the terminal
segment. The integumentary armature is almost entirely lacking in a great
many species but some, such as Stylops
californica (Fig. 218B),
bear numerous spines in a transverse ring on the posterior margins of the
abdominal segments. A few minute
setae are found on the dorsum of the head and thorax. In C.
antestiae (Fig. 218A), the
thoracic segments bear heavy plate‑like structures, with serrate
posterior margins, ventrally between the legs. The tracheal system has not been extensively studied in any
species, and the triungulinid is thought to be without spiracles. Nassanov as a simple apodous form, tapering toward both ends,
with a thin integument and no fleshy protuberances or processes, describes
the second‑instar larva of X. vesparum. Brues's description of that of S. melittae
noted a median row of 12 fleshy protuberances, considered to be locomotors in
function, on the venter of the body.
In Corioxenos antestiae, small paired
protuberances representing the legs are present on the thoracic segments. In the third instar of Corioxenos,
the male still retains traces of the legs, but these are lacking in the
female. The latter now shows the
first indications of median dorsal protuberances on the body segments. The fourth instar is quite similar, with
the leg rudiments conspicuous in the male, and the dorsal prominences of the
female are distinct. The latter are
conspicuous on the fifth instar of both sexes. In apparently all species, the sexes can be differentiated in
the third instar by the presence, in the female, of two or three columns of
massed cells extending the length of the abdomen on each side of the
intestine. The sixth‑instar female larva shows the differentiation of
the cephalothorax, which is as yet unsclerotized, with a pronounced
constriction at its juncture with the abdomen, and is deflected
ventrally. The spiracles of both
sexes and the mouth parts of the female are sclerotized. The seventh‑instar larva of both sexes of Corioxenos retains the fleshy
dorsal processes on the abdominal segments, though they are much larger in
the female (Fig. 219). In the male,
there are sclerotized bands on the cephalothorax, and after extrusion the
entire structure in both sexes becomes heavily sclerotized and brown in
color. The side in contact with the
host body, which in this species is the venter, is appreciably flattened in
the female. The cephalothorax of the male is almost quadrangular in form and
larger than that of the female. This
greater size of the male is also found in the species attacking Homoptera, as
contrasted with the opposite condition in the species attacking Hymenoptera. The fleshy dorsal processes disappear as
the body becomes distended with eggs.
These processes are considered to serve principally to increase the
absorptive area of the body, but they also function in a limited way in
locomotion. In all species, the
cephalothorax becomes heavily sclerotized, the color being white at the time
of extrusion but changing in a few hours to yellow, cinnamon, or brown. In certain species, the segmentation of
the cephalothorax is indicated at the lateral margins, and in Corioxenos distinct lines
reveal it in the male and definite constrictions in the female. Throughout the Strepsiptera the exerted
cephalothorax of the two sexes can be readily distinguished, that of the
female being appreciably flattened dorsoventrally, whereas that of the male
is almost cylindrical, its depth very nearly equaling the width. The Male Pupa.‑‑The male pupa is distinctive in form
and is found within the abdominal portion of the unbroken larval exuviae.
According to Pierce, the seventh instar is really a pupa, which is enclosed
by the sixth larval exuviae, and a second pupal instar then follows. After transformation, the adult male is
thus enveloped in three skins. In Corioxenos, the pupal skin is
pushed downward to the posterior end of the puparium. The Adult Female.‑‑Because of the retention of the
larval form by the adult females of the order, their persistence within the
host body, and the peculiar development of the reproductive system, it seems
desirable to give here a general description of that stage. It has been mentioned that in the apodous
forms the seventh‑instar female larva transforms directly into the
adult without the intervention of a pupal stage. Much controversy arose among early entomologists as to the
orientation of the body of the female, some claiming that the exserted
portion was the head and others that it was the posterior portion of the
abdomen. The latter appeared logical
in view of the emergence of the triungulinids at that point. There was also considerable doubt as to
the sides of the cephalothorax that were to be designated as dorsal and
ventral. The true interpretation was
arrived at only by an examination of the nervous system. The
adult female, as usually described, consists of the cephalothorax of the last
larval instar plus the abdomen of the adult.
Whether or not exuviation is complete is not known; if it does occur,
the head and thorax of the adult female must be undifferentiated, with the
cuticle thin, transparent, and following closely the inner surface of the
cephalothorax. Certainly, the
spiracles of the last larval instar, as well as other structures of the
cephalothorax, serve the adult, als0.
The exuviae of the abdomen remains closely enveloping the adult except
for a definite area ventrally, which draws somewhat away from the body to
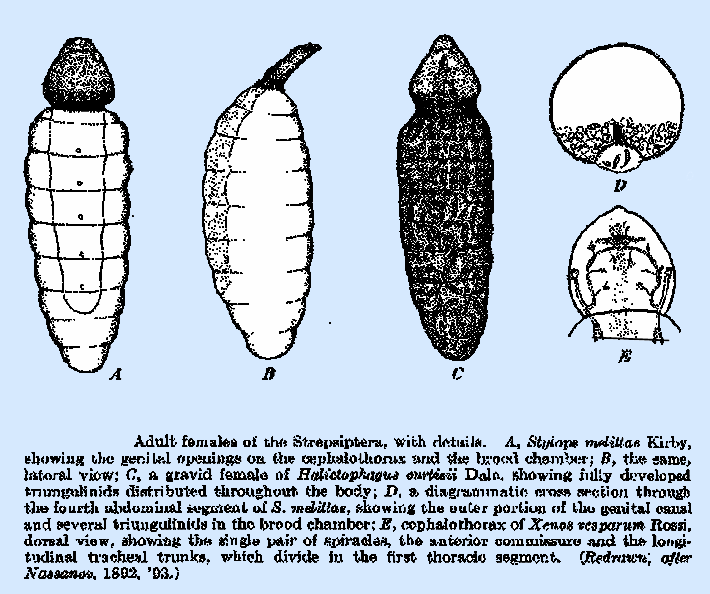
form the brood chamber (Fig. 220A).
According to Muir, the last larval exuviae of Elenchoides perkinsi
is broken over the dorsum of the abdomen as a result of the great distention
of the body of the female that accompanies egg production. According to Nassanov, the adult female of
S. melittae is enveloped in two larval exuviae rather than
only one. In many species, the portion of the exuviae forming the brood
chamber becomes considerably darkened, in contrast to the white of the
remainder of the body. The brood
chamber varies in width and length among the different species and extends to
the posterior margin of the fourth segment in Dacyrtocara undata
and to the eighth in Xenos vesparum, while in other
species it is of intermediate length.
This depends on the number and position of the genital pores. The chamber usually extends one, and
occasionally two, segments beyond the last pore. The epithelium lining the ventral surface of the body and
forming the dorsal wall of the brood chamber is appreciably modified, with
the cells produced into spiny processes (Fig. 220D). This spiny surface is said to facilitate
the escape of the triungullinids. Westwood originally called attention to the existence of a cord
or tube connecting the base of the abdomen to the integument of the host, and
it was suggested that food is taken into the body by this means. Pierce observed the same feature in Halictoxenos crawfordi Pierce and in several
Xenidae, but no definite information is available regarding either its
structure or function. Ulrich stated
that it is of host origin and that it serves as a means of attachment. It is quite possible that this structure
represents an ingrowth of the integument, or rather a scab formation,
analogous to the respiratory funnel formed about the posterior end of parasitic
tachinid larvae that maintain, for respiratory purposes, a constant
communication with the exterior through an aperture in the body wall of the
host. The wound resulting from the
extrusion of the cephalothorax is large, and, without complete healing,
resulting in a close union of the intersegmental membrane with the
"neck" of the parasite, the host would soon die from desiccation or
disease. The characters that have been utilized for describing the
larviform females are as follows (Pierce, 19O9, 1918): Measurements: Head--width at emargination at base of mandibles in
relation to thorax. Cephalothorax--width at base of head, at
spiracles, and at base; length from front edge of spiracle to apex of head and from base to apex of
cephalothorax. Mandibles--presence or absence, mld forms. Spiracles--number, position, and form. Genital pores--number and position. Genital openings--number and form. Distinct mandibles are present in the heads of the adult females
of the Stylopidae, Xenidae, and Hylecthridae. In a few species, these are
simple and small, but in the majority they bear a distinct tooth at the inner
apical angle and may be widely separated.
They are vestigial or absent in the Halictophagidae and
Elenchidae. These widely spaced thorn‑like
mandibles may be utilized by the mature larva in effecting the extrusion of
the cephalothorax from the body. The tracheal system of the mature larva usually comprises a pair
of large spiracles on the cephalothorax, with branches extending throughout the
body. In X. vesparum
(Fig. 22OE) and S. melittae, according to
Nassanov, the two main anterior trunks are directed caudad from the
spiracles; just before leaving the cephalothorax, each one gives off a branch
that extends forward, and the two then unite to form a dorsal commissure. The main trunks divide immediately after
entering the abdomen and reunite in the eighth segment, thus giving a dorsal
and ventral trunk, with various branches, on each side of the body, and a
posterior commissure. Smith and Hamm,
however, noted only a single trunk on each side, with numerous ramifications,
in S. melittae.
According to Pierce, the paired lateral trunks are normal for the
order. With few exceptions, the adult
females have a single pair of rather large spiracles situated at the lateral
margins of the metathorax. Callipharixenos muiri and Chrysocorixenos siamensis
Pierce have two pairs, and D.
undata is distinguished by
three pairs of small, slit‑like spiracles at the margins of the
abdominal brood chamber, in addition to the thoracic pair. Presumably, the metathoracic spiracles are
closed until the extrusion of the cephalothorax from the body of the host. The exact stage in larval development at
which the spiracles appear is unclear, though Nassanov and others have
implied that they occur in all instars after the first. The
reproductive system of the female is of remarkable form and not at
all comparable with that found in any other insect. The eggs lie free in the body cavity,
though Brues detected a rudimentary oviduct and vagina in Xenos wheeleri. On the
median ventral line of the abdomen, there first appears a series of
invaginations of the cuticle that develop into funnel‑like structures,
the inner ends of which become open about the time the larvae are
mature. These are the genital duct or
pores, through which insemination takes place and the larvae later
escape. These pores are usually found
on the second and following abdominal segments, though in Corioxenos antestiae the first one is on the third segment. The number of pores is two in the case of Dacyrtocara undata; three in Elenchinus japonicus E. & H., Elenchoides
perkinsi and Carwfordia labiata Ogl.; four in X.
bohlsi, X. vesparum,
(Fig. 221), X. wheeleri, Tridactylophagus mysorensis,
and Corioxenos antestiae; and five in Halictoxenos viridulae Pierce, Halictophagus curtisii Dale, Chrysocorixenos siamensis, Callipharixenos muiri,
and several species of Stylops. A very unusunl arrangement of these pores
is found in Stichotrema dallatorreanum, in which
species they occur in three transverse rows of 12-14 each on the first or
second abdominal segment. Hofeneder has erected the superfamily
Stichotrematoidea on the basis of this character. The genital opening usually occurs as a transverse crescentic
slit ventrally at the juncture of the head and thorax; it is through this
opening that insemination takes place and the triungulinids later
emerge. There is considerable
variation in the form of this opening among the different species; it is
almost semicircular in Elenchoides
and Elenchinus. The opening is usually closed by a
membrane, which is perforated at the time of mating. A striking departure in respect to the
genital opening is found in Corioxenos
antestiae in which, instead
of being single and medially placed, it is paired, with one opening occurring
at each side of the cephalothorax.
This arrangement is said to be essential in order that mating may be
accomplished, for the cephalothorax, except for one lateral margin, is
covered by the wing of the host. The free‑living
adult females of Eoxenos
(Fig. 217B) and Mengenilla
differ in many morphological characters from those which are permanently
internal in habit. They are wingless
and have primitive compound eyes, and the single genital opening occurs
ventrally at the posterior margin of the seventh abdominal segment. In Eoxenos, the antennae are five‑jointed
and the legs have four‑jointed tarsi terminating in a pair of claws,
whereas in Mengenilla they
are four‑ and three‑jointed, respectively. The
reproductive system of E.
laboulbenei, which has been
studied by Parker and Smith, is quite different from that already described
and approaches the type common to that of most other insects. The vagina is distinct and terminates
anteriorly in the fifth abdominal segment, at which end it is apparently
open. The eggs lie free in the body
cavity, and some may even be contained in the head. There is no evidence of segmental genital pores on the median
ventral line of the abdomen.
Spiracles are present on the first seven abdominal segments. References: Please refer to <biology.ref.htm> [Additional references may be found at: MELVYL Library] |