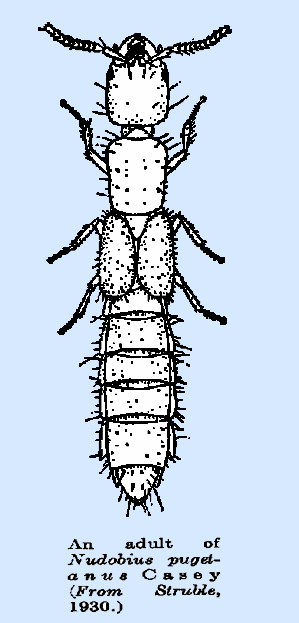
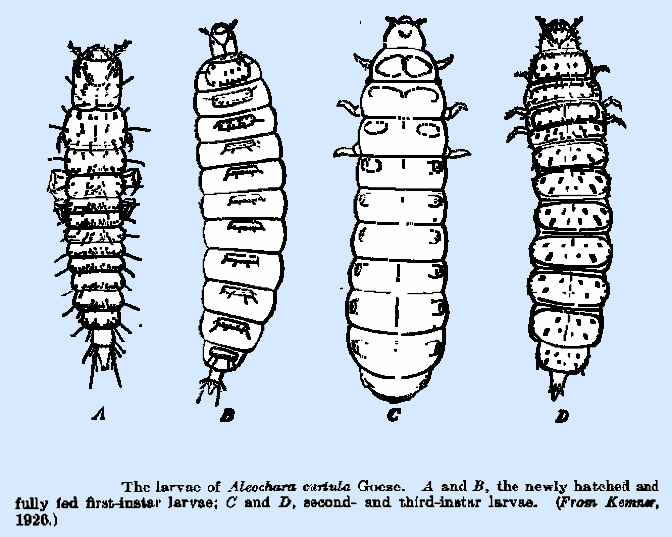
|
File: <satph1.ima.htm> [For educational purposes only] Terminology Glossary <Principal Natural
Enemy Groups > <Citations> |
|
Immature
Stages of Staphylinidae
First instar larvae of Staphylinidae are not easily distinguished
from those of the predaceous and scavenging species. These larvae first search in the soil to a
depth of 2-15 cm until a Hylemya
puparium is found, and it then penetrates the hardened shell. The puncture is sealed after entry. The body of newly hatched larvae is ca.
1.5mm in length, rather slender, and distinctly segmented, with 10 abdominal
segments, of which the last two are darker in color and taper to a rounded
point. The head is large and darker
in color than the body, and the antennae are 3-jointed and well
developed. Legs are normal for the
family. The caudal cerci are shorter
than those of most other species and are borne on short cylindrical processes
dorsolaterally at the posterior margin of the penultimate segment. Following entry of the host puparium, the larva begins feeding
through a minute puncture made in the delicate cuticle of the pupa. The feeding position is frequently changed
but apparently limited to the anterior dorsal region. When fully fed the body is considerably
enlarged, the transparent intersegmental membranes being greatly stretched
and the average length is ca. 2.0 mm (Clausen 1940/1962). Second instar larvae are markedly
different from the first, being very degenerate as a result of the adoption
of a parasitic mode of life. The body
is glistening white, with the cuticle very lightly sclerotized, and the setae
and cerci are absent. The legs are
rudimentary and indistinctly segmented and lack the large terminal claw. Each of the last two thoracic and the
first eight abdominal segments has a single pair of fleshy prominences at the
dorsolateral margins. Feeding by this
stage is extensive and very little movement occurs (Clausen 1940/1962). Third instar larvae are generally
identical to second, but larger.
There are also 9 pairs of spiracles situated between the first and
second thoracic segments and on the first eight abdominal segments,
respectively. in Aleochara and other genera that pupate outside the host
puparium, the segmental tubercles or prominences are lacking. During the feeding period no excrement is
voided with the exception of occasional minute drops of a clear fluid, and
the mature larva casts the meconium.
The host pupa is completely consumed, and the puparium becomes opaque
because of the meconial covering on the inner surface. Please CLICK on picture to view
details:
In summer the feeding period is complete in 5-6 days after the
first molt and is followed by a quiescent stage of 12-18 days prior to the
appearance of the pupa. The pupal
stage also requires 12-18 days at the completion of which the adult beetle
gnaws its way out of the puparium (Clausen 1940/1962). Adults are very active and feed
extensively on cabbage maggots. There
are apparently two generations/yr, corresponding to the cycle of the host,
and the winter season is passed as a first-instar larva within the host
puparium. The first brood of adults
appears in May and June and the second in August and September. Although most parasitic members of the family attack puparia
occurring on or in the soil, occasional species find their hosts on or in
plants. Maseochara valida
Lec. was found by Coquillett (1891) to develop in the puparia of a syrphid
fly, Copestylum marginatum Say, which develops
in the semiliquid material in the decaying leaves of cactus. Kramer (1926) studying Aleochara curtula Goeze, parasitic in the puparia of Lucilia, etc., found that this
species habits are generally similar to those of C. bilineata,
except that the third instar larva assumes an active and more normal form, with
the legs being large and well developed for locomotion; and it emerges from
the puparium for pupation in a cell in the soil. This habit is found in the genera Aleochara, Baryodma,
Polychara, and Maseochara, whereas the more
degenerate form, which pupates within the host puparium, is found in Coprochara and Polystoma. Baryodma bimaculata Grav. was noted by
Lindquist (1936) to develop in the puparia of Sarcophaga and Cryptolucilia
in Texas. The planidium enters the
puparium in the same way as Coprochara,
and emergence of the adult occurs ca. 20 days later. The field parasitization of these hosts is
ca. 25%. Observations recorded on other species of parasitic Staphylinidae
suggest that the adults of these species are also important as enemies of the
same pests because they are predaceous on both larvae and pupae (Clausen
1940/1962). Quayle (1913) recorded
the habits of Somatium oviformis, a minute species
that seems to feed mainly on red mites.
This is true of both the adult and larval stages. The eggs are light orange in color and are
laid singly on the undersides of the infested leaves. Larvae consume ca. 20 mites/day and adults
about half as much. Mank gave an
account of the habits and descriptions of the immature forms of a series of
species predaceous on dipterous larvae occurring in decaying vegetable
matter. These species belong to the
well known genus Philonthus
and related groups. Both adults and
larvae feed extensively on maggots found in the medium in which they
live. The life cycles of the
different species are found to be relatively short (one month or less). The larvae of predaceous species are very
active and aggressive. The body is
elongate in form, and individuals may be readily distinguished from carabid
larvae by the prominent two-jointed caudal stylets and by the single claw of
the tarsi. They also generally lack
the distinct heavily sclerotized segmental plates often found in Carabidae,
although these plates are present in Tachinus. The most reliable character in distinguishing
larvae of the family is the "upper lip," which varies in the number
and size of the teeth borne at the anterior margin. The abdomen terminates in a relatively large
"pseudopod" or "pusher," which is utilized in
locomotion. Clausen (1940) stated that the pupae present few distinguishing
features, with the main character utilized is the fringe of hairs at the
anterior margin of the thorax and those at the lateral margin of the
abdomen. In Tachinus, the integument of the pupa is soft and the body
is completely covered with a soft material which becomes silvery in
appearance. White & Legner (1966) give a detailed account of the biology
of Aleochara taeniata Erichson, attacking
muscoid flies. Legner & Warkentin
(1991) considered species of Philonthus
very important predators of field breeding Muscidae. This parasitoid/predator was introduced in
California from Jamaica. Adults are
voracious predators of house fly eggs and young larvae. Eggs hatch in ca. 4 days and larvae search
actively through the substrate for fly pupae, the parasitoid-susceptible
stage. Pupae are entered through a
hole gnawed in the puparium wall by the young larvae; the hole is closed with
what seems to be fecal matrial. The
three larval instars are ectoparasitic on the fly pupae within the
puparium. Mature larvae emerge into
the substrate where pupation occurs.
Larval development requires 6-7 days, the pupal development another
14-16 days at 23.9°C. The total life
cycle at this temperature is ca. 25 days from egg to adult. Also please
see separate Staphylinidae section. References: Please refer to <biology.ref.htm>, [Additional references may be found at: MELVYL Library] |