|
čFile: <sceli1.ima.htm> [For educational purposes only] Terminology Glossary <Principal Natural
Enemy Groups > <Citations> |
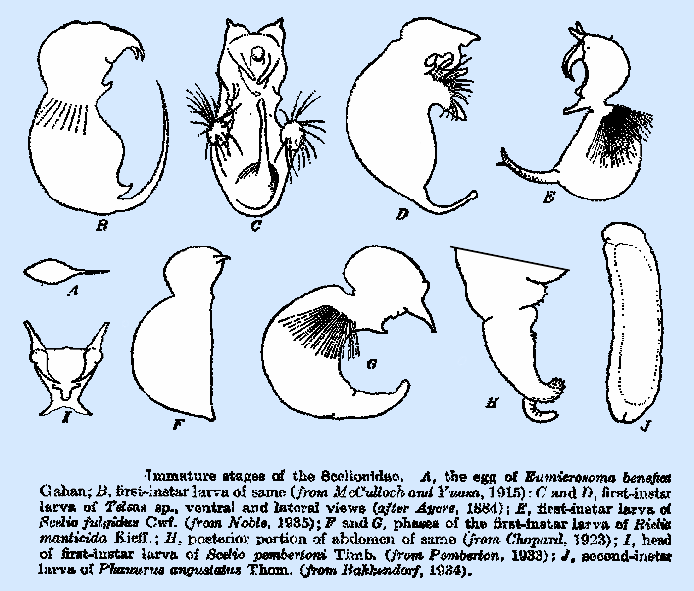
Immature Stages of Scelionidae
Immature stages of Scelionidae were discussed in detail by
Clausen (1940), as follows: The eggs of the species
of Scelionidae that have been described are of a uniform type, all being
stalked, with the main body ovate to spindle shaped and the tapering or
tubular anterior stalk ranging in length from 1/2 to 1 & 1/2 X that of
the main body. The eggs of Scelio are slender, with the
line of demarcation between the stalk and main body not distinct, and that of
S. pembertoni has a small pedicel at the posterior end,
also. There is an increase in size
during incubation, as a result of which the stalk disappears. The
first‑instar larva of the
family is "teleaform," so‑called from the larva of Tiphodytes (Teleas sp.) described and
figured by Ganin (1869). It is
characterized by a complete lack of segmentation but with the body divided by
a sharp constriction into two almost equal portions. There is a difference of opinion as to the
parts that constitute the anterior portion.
Henriksen, Bakkendorf (1934), and Pagden (1934) considered that it
represents the head alone, while Noble & Kamal termed it the
cephalothorax, made up of the head and the three thoracic segments. This latter interpretation is more
probably correct. The mandibles of
all species are external, widely spaced, exceedingly large, curved, and
sharply pointed and may be either heavily sclerotized or fleshy and
unsclerotized. In Phanurus sp. dissected from
eggs of Chrysopa in Japan,
there are no other evident structures or organs on the cephalothorax, while
in several other species of the family various head structures are well
developed. The antennal processes of Scelio fulgidus (Fig. 113E) and S. pembertoni
are large and conical, are widely spaced, and arise immediately above the
bases of the mandibles. In a number
of species, there is a large, fleshy lobe or process on the median ventral
line of the cephalothorax, below or behind the mandibles. This is highly developed in the genus Scelio (Fig. 113I) and has been
considered the labium by several authors. Please CLICK on
picture to view details:
The
abdomen is almost globular in form and terminates in a caudoventral
horn, or tail, which may be fleshy and of irregular form or heavily
sclerotized, sickle like, and terminating in a sharp point. In some species, there are one or two
supplementary lobes at the base of the tail. The fleshy type of tail is usually spined dorsally and on the
distal portion and is occasionally bifurcate. at the tip. In Eumicrosoma, Telenomus, Scelio, Phanurus,
and Microphanurus, and
probably in other genera also, there is a partial or complete transverse ring
of long hairs near the anterior margin of the abdomen. These hairs vary considerably in number
and distribution. In E. benefica (Fig.
113B), they occur upon the sides only, whereas in others the ring is
complete and it is double in several species of Phanurus. The abdominal
hairs of Teleas sp. (Fig.
113C, D) figured by Ayers (1884), Limnodytes,
and Tiphodytes are in
distinct tufts upon the summits of a pair of fleshy lobes situated at the
lateroventral margins on the anterior portion of the abdomen. Marchal (l900) illustrated them in that
arrangement in T. gerriphagus Mnrchal, who;e
Martin (1928), dealing with the same species, shows the hairs in a transverse
row. Chopard (1923) figured several supposed developmental phases of
the first‑instar larva of Rielia
manticida in the eggs of
Mantidae. The first (Fig. 113F),
secured from host eggs in April, is of simple form, with the abdomen lacking
the band of hairs and the tail. Those
found in May show a lateroventral tuft of short hairs, and the tip of the
abdomen is produced into a broadly conical tail. The form found in June and July (Fig. 113G) has the lateral
abdominal hairs well‑developed and the tail further enlarged (Fig. 113H).
The author is inclined to consider the latter to be the second
instar. It is however, identical in
general characters with the first‑instar larva of various other
species. The
second‑instar larva had been
described in only a few species as of 1940 (Clausen 1940). That of T. gerriphagus
figured by Martin is irregularly ovoid in form, with the mandibles still
large and a small hook‑like caudal horn. Immediately above the mandibles are two plate like thickenings
of the integument, separated by a median depression. The abdominal hairs are present in groups
of 5-6 in a band across the dorsum and sides. None of these characters was found in all of the specimens
examined, and the true form of the second instar is thus doubtful. That of M. basalis
is very robust, with the segmentation indistinct; the mandibles are small and
simple. There are no integumentary
spines or setae, and the caudal horn is lacking. The second‑instar larvae of T. ulyetti and Phanurus angustatus Thom. (Fig. 113J) are cylindrical and
distinctly segmented, but otherwise similar to that of M. basalis.. Neither the first‑ nor second‑instar
larva of any species has been found to possess a tracheal system or
spiracles. The
third‑instar larva of T. ulyetti is similar in form to the second but may be
readily distinguished by the presence of nine pairs of spiracles, situated on
the last two thoracic and the first seven abdominal segments. Kamal mentioned that only the two pairs of
thoracic spiracles are functional in M.
basalis, the following seven
being minute and closed. The
integument of the abdomen bears numerous small tubercles at the lateroventral
margins, which extend across the venter on the posterior segments. This species and S. fulgidus
are grayish‑green in color. The
mature larva of Cacus oecanthi Riley bears a pair of
rounded protuberances Internally on each body segment except the last, and
the second to seventh segments also have a pair of prominent tubercles
dorsally (Parrott & Fulton 1914). In n number of species, only two larval instars are mentioned,
this being due presumably to the marked similarity of the second and
third. The larva of P. angustatus (described and figured as the second and last
instar (Bakkendorf 1934) is probably the true second, as judged by the lack
of a respiratory system. References: Please refer to <biology.ref.htm> [Additional references may be found at: MELVYL Library] |