|
čFile: <rhiph1.ima.htm> [For educational purposes only] Terminology Glossary <Principal Natural
Enemy Groups > <Citations> |
Immature
Stages of Rhipiphoridae
Some detailed information on immature stages of Rhipiphoridae = Ripiphoridae)
was given by Clausen (1940) The eggs of the various species are relatively minute, ranging
from 0.3 mm. in length in Macrosaigon
flabellatum to 0.7 by 0.15
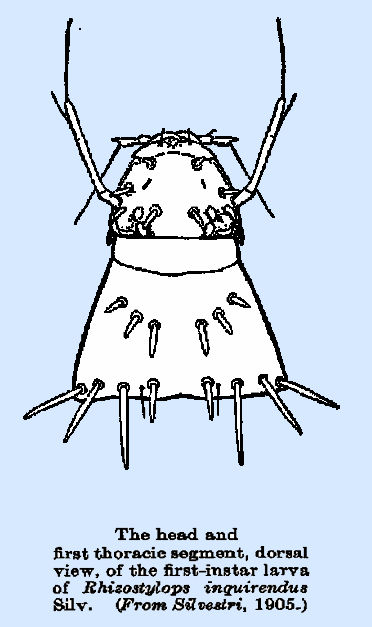
mm. in Rhizostylops inquirendus. They are usually broader at one end,
white, and in Meteocus paradoxus, at least, covered
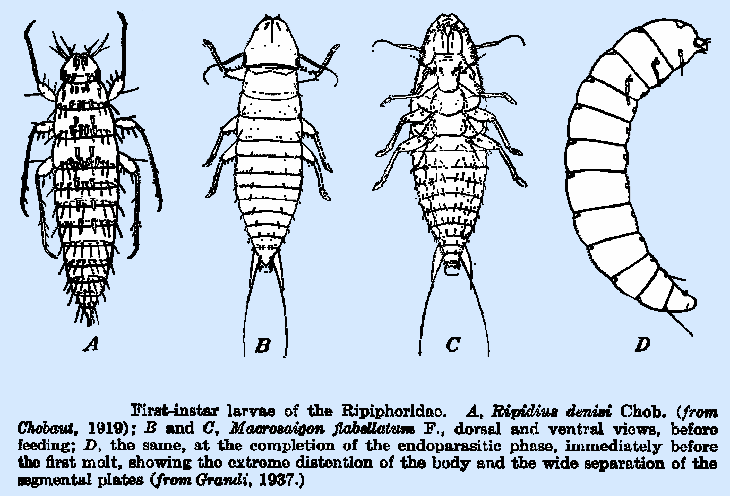
with a mucilaginous material. The triungulinid first‑instar larvae are of the planidium
type, with the body smoothly tapered and the segments each bearing a heavily
sclerotized band, with pleural plates, and separated by thin and very elastic
intersegmental membranes, which are visible only after the commencement of
feeding. The body color is blackish,
owing to the heavily indurate segmental plates. The thoracic segments are large and broad, and the abdominal
segments, which apparently range in number from 8 to lO, are of diminishing
length. The head (Fig. 235) is
somewhat triangular in form and heavily sclerotized and bears several ocelli,
placed close together, at each side.
The antennae are usually two‑jointed, the second terminating in
a long hair‑like process that is believed to be a segment. The mandibles are heavy, simple, and
sharply curved. The legs are long and
slender, and the tibiae usually terminate in large leaf‑like
pulvilli. The body segments bear a
variable number of integumentary setae and spines. In the majority of species, the two caudal segments each bear a
pair of cerci of varying length, and thc abdomen terminates in a suctorial
disk. Please CLICK on
pictures to view details:
The characters of the triungulinid, which show marked variation
between species, are: the antennae, the claws or pulvilli of the legs, the
setae and conical spines on the body segments, and the caudal cerci. In most species, the antennae are two‑jointed,
the first often bearing a spine or branch near the distal end, though that of
Macrosiagon tricuspidata is believed to be
three‑jointed. The large leaf‑like
pulvillus that is borne at the end of the tibia, in lieu of the tarsus, is
replaced in R inquirendus by a pair of minute
claws, whereas in Ripidius denisi (Chobaut 1919) the claws
are present and also a small pulvillus.
In M. flabellatum, pleural plates are
prcsent on the sccond and third thoracic and the first seven abdominal
segments (Grandi, 1937). Perhaps the
most conspicuous specific character is the number and arrangement of the
spines and setae upon the different body segments. A number of species, including M tricuspidata,
M. flabellatum (Fig. 234B,C), and Ripiphorus solidaginis,
apparently have only a single seta at each lateral margin of the segments,
and Metoecus paradoxus has a row of small
setae dorsally and ventrally on each segment. Ripidius denisi and Rhizostylops inquirendus
are distinguished by long setae at the posterior margins of the segments and
heavy conical spines on the head and thorax, a pair of these occurring also
on each of the first three abdominal segments in the first‑named
species. With respect to the caudal
cerci, Ripiphorus solidaginis has only two very
short pairs, whereas Ripidius
denisi has one pair, these
being only slightly longer than one or two segments. M.
paradoxus and Macrosiagon cucullatum have a single long
pair, also, but these are equal to the body in length. Other species have two pairs of unequal
length; those on the last segment are longest and equal or exceed the body
length in M. tricuspidata and M. flabellatum. In
the latter species, the first pair occurs on the ventral plate of the
penultimate segment, and the second pair is on the dorsal plate of the last
segment. In Rhizostylops inquirendus and M. flabgellatum, the spiracles occur only on the last two abdominal segments, and transverse
commissures occur at the caudal end of the body and in the second and third
thoracic segment, whereas there are nine pairs in the case of M. flabellatum, these being situated on the pleural plates of
the mesothoracic and the first seven abdominal segments and at the lateral
margins of the dorsal plate of the eighth segment. Little is known of the second‑instar larva of any species.
Stamm mentioned it in the case of Ripidius
pectinicornis stating that
it is quite similar to the mature form.
It is to be expected that all the distinguishing characters of the
triungulinid would be lost at the first molt; and the second instar, being
strictly a feeding form, is grub‑like.
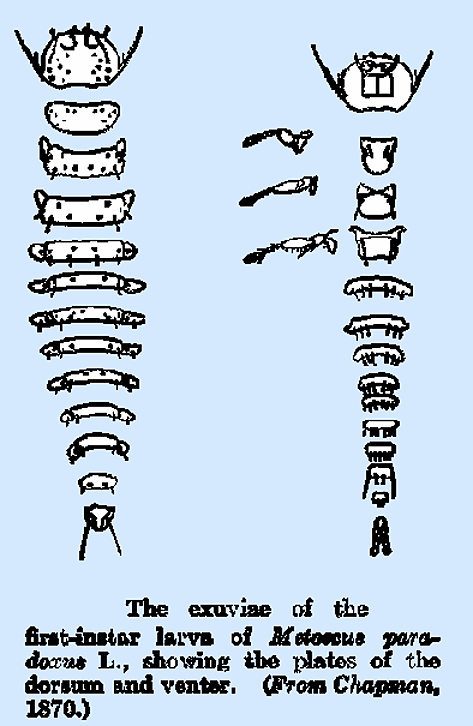
What is presumably the second instar of Metoecus paradoxus
was figured by Chapman; it is white in color and distinctly segmented, the
legs are entirely lacking, and the tracheal system has spiracles on the first
and third thoracic and the first six abdominal segments and accessory
longitudinal trunks in the thorax. The mature, and presumably third‑instar, larva is known in Macrosiagon flabellatum, Metoecus paradoxus, Ripiphorus
stylopides, and Ripidius pectinicornis.
These present considerable differences, the last‑named being
markedly different from the others.
In Macrosiagon flabellatum (Fig. 233A), the
body is rather elongated and greatly curved, with large fleshy processes on
all segments, and the legs are short, fleshy, and nonfunctional. Open spiracles occur on the mesothorax and
on the first seven abdominal segments, and atrophicd spiracles are found on
the metathorax and on the eighth abdominal segment. The larva of Metoecus
paradoxus is also rather
elongated, with the head and thorax sharply bent ventrally and the caudal
segments tapering and likewise curved ventrally. A number of fleshy tubercles occur upon the thoracic segments,
and the legs are rudimentary, being conical, somewhat pointed, and divided by
mere constrictions into three parts.
There are eight pairs of spiraclcs, situated as in the second
instar. The mature larva of Ripiphorus stylopides was figured by Böving and Criaghead (1930) and
presents several features that distinguish it from M. paradoxus. Two pairs of large fleshy conical
processes occur dorsally and dorsolatcrally on each body segment, and the
eight pairs of spiracles are situated as in Macrosiagon flabellatum. In contrast to the larvae already described, that of Ripidius pectinicornis (Fig. 237) may be considered as intermediate
between them and the normal coleopterous larva. The fleshy segmental processes are lacking, and the legs (Fig.
237c) are larger and distinctly four‑segmented and terminate in a
heavily‑sclerotized claw like tarsus.
The head and mouth structures are much reduced (Fig. 237a, b), the
antennae being short and two-jointed, and the most conspicuous feature is
the labium which ends in three- jointed palpi. No mandibles are discernible.
Dorsally on the head there is found a pair of curved, converging,
heavily chitinized ridges, which are present also, though widely separated,
on the thoracic segments. They are
believed to serve in perforating the integument of the host at the time of
emergence from the body. Such an
adaptation is not required in the species that develop externally upon hymenopterous
larvae. References: Please refer to <biology.ref.htm> [Additional references may be found at: MELVYL Library] |