|
File: <ptero1.ima.htm> [For educational purposes only] Terminology Glossary <Principal Natural Enemy
Groups > <Citations> |
Immature
Stages of Pteromalidae
Immature stages of Pteromalidae were discussed in detail by
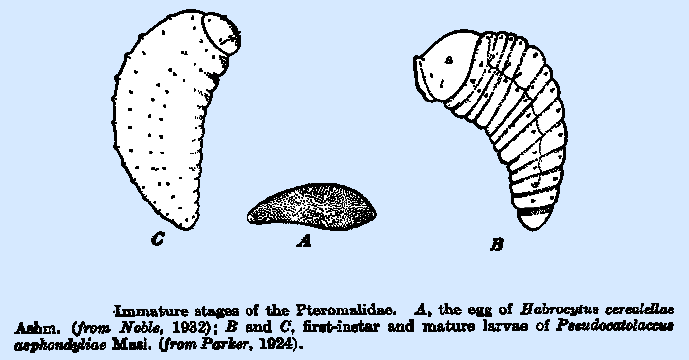
Clausen (1940), as follows: The eggs of the great majority of the species of the Pteromalidae
are ovate, ellipsoidal, or cylindrical in form, often slightly curved, and
without an anterior stalk or pedicel of any sort. That of Enargopelle
ovivora, however, is an
exception in that it has a slender stalk, one‑third as long as the egg
body, at the broader end. In many
species, the chorion is smooth and shining; in others, it is covered with
minute spines or spicules, but with the poles bare (Fig. 55A). These spicules often give the egg a
grayish color. The eggs of Dibrachys cavus, Stenomalus
muscarum, and Habrocytus trypetae Thoms. are covered with "tubercles," or
papillae; those of Pachyneuron
coccorum L. have a granulate
surface. In Dibrachoides dynastes,
the chorion bears longitudinal ridges.
The sculpturing of the egg is not uniform even within a genus, as
shown by the smooth glistening egg of Pteromalus
puparum and the spinose
chorion of that of P. variabilis
Ratz. The sculptured chorion is found
only upon eggs that are deposited externally. Please CLICK on picture to view
details:
The larvae of practically all species are hymenopteriform with 13
distinct body segments, the head often large, and the integument bare except
for three pairs of setae on each of the thoracic segments and two pairs on
those of the abdomen, the four pairs of spiracles are situated on the second
thoracic and the first three abdominal segments (Fig. 55B). There appears to be a variation among the
species in the number of sensory setae.
The body of Stenomalus
micans is covered with
minute integumentary setae, and the last abdominal segment is modified to
form a "furca" which serves to hold the larva in a favored feeding
position. Habrocytus sp. reared from braconid cocoons by
Voukassovitch (1927) has a sucker‑like organ ventrally on the second
thoracic segment, which is stated to serve a locomotory function. Merisoporus
chalcidiphagus W. & R.
has an additional pair of spiracles, which is on the third thoracic
segment. Certain of the endophagous
species, such as Pteromalus puparum, lack an open tracheal
system. The larva of Pirene
graminea, described and
figured by Kutter (1934), departs from the normal of the family and Is
distinctly mandibulate in form. The
head is large, the body segmentation indistinct, the integument without
setae, and the large, extruded, falcate mandibles are very widely spaced and
lie transversely. Marchal (1907)
described a larva of similar form in Tridymus
piricolaa Marchal. The full complement of five larval instars has been described for
Eupteromalus nidulans, E. fulvipes
Forbes, Dibrachoides dynastes, Pachycrepoideus dubius
Ashm., S. micans, and Merisus destructor Say. Dibrachys cavus and H.
cerealellae, both of which
have been studied in detail, apparently have only four, and Habrocytus sp. discussed by
Dustan has only three. Kutter
described only two for Pirene
graminea, though his prepupa
shows distinctive characters representing presumably a third instar. The second‑instar larvae of all species are of simple form,
with the sensory and integumentary setae reduced in size; the latter are
often absent. The nine pairs of
spiracles occur on the last two thoracic and the first seven abdominal
segments. In H. cerealellae, only the four
pairs that persist from the first instar are open immediately after the molt,
and the additional five, which are smaller, appear later in the stage. The larva of P. graminea
is indistinctly segmented, and the head is much reduced, with the mandibles
small, curved, and very widely spaced. The fourth‑instar larva of S. mican
is distinguished from other larvae of the family by a heavily sclerotized
boring armature on the head. This
consists of a transverse plate with a serrate edge situated immediately above
the labrum and one or two heavy conical spines on the median line below the
antennae. The mature larvae of the different species are uniform in their
characters and present no general distinguishing features. The mandibles are simple, and the integument
is smooth, with the sensory setae reduced in size. In H. cerealellae, there are three
pairs of prominent spines at the end of the last abdominal segment. This species and Pseudocactolaccus asphondyliae,
show pronounced intersegmental ridges dorsa11y. The tracheal system usually has nine pairs of spiracles,
situated as on the second instar.
However, Haviland (1922b) recorded 10 pairs for Asaphes vulgaris
Wlk.; the first of these is situated on the intersegmental membrane between
the first and second thoracic segments, and those following are on the third
thoracic and the first eight abdominal segments. There is a vestigial tenth pair on the eighth abdominal segment
in H. trypetae Thoms.
The spiracles of S. micans first appear on the
fourth instar, and rudimentary spiracular stalks are also present on the
first thoracic and the eighth abdominal segments. Kutter's figure of the prepupa of Pirene graminea,
which bears the external structures of the mature larva, shows the last
segment produced into tubelike form and bearing two pairs of strong setae at
the distal end. This tube is stated
to be retractile. Kearns describes an unusual development of the internal tracheal
system in the endophagous first‑ and second‑instar larvae of S. micans (Fig. 56).
Both instars possess the usual longitudinal trunks, with dorsal and
ventral commeasures at the anterior and posterior ends, respectively, and
blunt spiracular stalks in the last two thoracic and the first eight
abdominal segments. In addition, a
pair of visceral tracheal trunks arises from the anterior commissure and extends
over the dorsum of the intestine to the eighth abdominal segment, where they
unite. These visceral trunks were not
detected in the third‑ to fifth instar larvae. A comparable modification of the tracheal
system was not known in any other hymenopterous larva in 1940 (Clausen 1940).
Another departure from the normal respiratory system is described
by Dustan (1923) for the larva of Habrocytus
sp. parasitic in Rogas pupae. His
description of that of the mature larva is as follows: " Perhaps the
most amazing thing about this parasite is its immense tracheal system which,
as was said previously, almost fills the body cavity. It has a tracheal trunk running down each
side of the body and spiracles that can be made out under the 4 mm.
objective, which appear to be closed, however; but more unusual than all, it
possesses myriads of tracheids packed into every conceivable part of the
body. These tracheids are collected
into bundles or areas that are held in place by a definite wall or membrane. Just inside the wall of each bundle we
find a ring of tracheids, varying somewhat in size but alike in having
extremely thin walls. All the space
inside the tracheids is packed with blood corpuscles and plasma, so that
each bundle really consists of a tracheal sheath, the inside of which instead
of being hollow is filled full of blood....
These tracheids open at the hypodermal wall and in this way secure an
abundant supply of oxygen from the blood stream of the host. This oxygen is then carried in the
tracheal bundles to all parts of the parasite and distributed by the blood
stream to the different organs and tissues." Also please
see Gerling & Legner (1968) work on Spalangia cameroni. References: Please refer to <biology.ref.htm>, [Additional references may be found at: MELVYL Library] |