|
File: <meloi1.ima.htm> [For educational purposes only] Terminology Glossary <Principal Natural
Enemy Groups > <Citations> |
Immature
Stages of Meloidae
Clausen (1940) remarked that he eggs of the different species of
the Meloidae are quite consistent in form, being somewhat cylindrical and
three or four times longer than wide, and with both ends smoothly
rounded. The chorion is delicate and
unsculptured, and the color ranges from white to yellow and orange. The size of the egg is governed not only
by the size of the beetle itself but by the number produced by the particular
species. Thus, in a species that
produces 3,000 to 5,000 eggs, they are markedly smaller than in one that
deposits only a few hundred. The larvae of the Meloidae pass through a rather striking
metamorphosis in which six instars are usually recognizable. Riley (1883)
proposed that they be designated as follows: Triungulin‑‑first‑instar larva Caraboid (Carabidoid)‑‑second‑instar larva Scarabaeoid‑‑third‑ and fourth‑instar
larvae Coarctate‑‑fifth‑instnr Inrva Scolytoid‑‑sixth‑instar larva These distinctive forms are readily recognizable in all species
which develop in locust‑egg masses, but the later instars are not so
well‑differentiated in those which attack bees. Among the latter, the coarctate larva has
been variously termed the pseudolarva, pseudonymph, pseudopupa, and
pseudochrysalis. The first‑instar larva has attracted the interest of
entomologists for a long period of time, and for many years its identity was
not established. It was at first
considered to be an adult insect, externally parasitic upon bees; it was
named Pediculus apis by Linnaeus, and later the
genus Triungulinus was
erected for it by Dufour. The term
"triungulin," later modified to "triungulinid," has
consequently been used to designate the larvae of the family; the reference
is to the three claws borne at the end of the tibia in the forms
described. This character is not
constant throughout the family as pointed out by Cros (1917) and Böving
(1924), there being a wide variation among the different species, and the
term is not applicable to a considerable portion of the family. Several later authors have used the term
"primary larva" instead, but this is likely to lead to confusion,
for it generally indicates a host‑parasite relationship. In all essential respects, this larva is
of the true planidium type, having the distinctive features and habits of
larvae of the Perilampidae, Eucharidae, Cyrtidae, Ripiphoridae, and
Strepsiptera. Please CLICK on picture to view details:
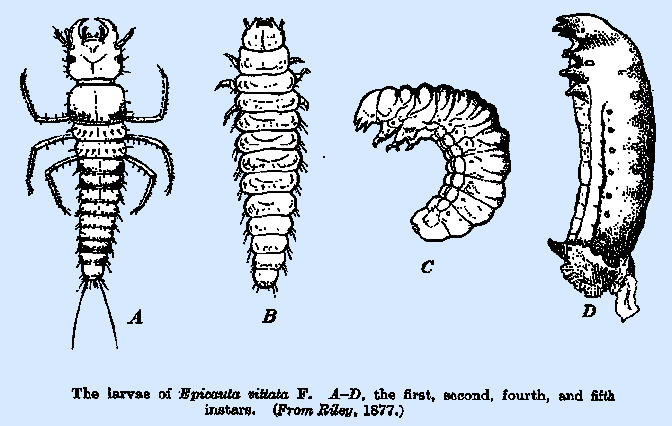
The body of the first‑instar larva (Figs. 239A, 240A) is
fusiform, is broadest in the thoracic region, and has the nine abdominal
segments successively narrower. It is
white, yellow, or orange at the time of hatching; but the integument quickly
darkens, often becoming almost black.
The head and thorax combined are approximately equal to the abdomen in
length. In outline, the head varies
from triangular to quadrangular. The
black ocelli, either simple or double, are situated nenr the lateral
margins. The mandibles range from the
simple falcate form to those having up to 12 teeth, the more strongly dentate
forms being of species that attack locust egg masses. The antennae are large, cylindricnl, and
three‑jointed; the second joint is longest and bears a sensory organ,
and the third terminates in a spine that may vary in length from half to six
times that of the entire antenna. The
thoracic segments arc long and heavily sclerotized, and the legs comprise
four principal joints, with the tibia terminating in a simple claw‑like
tarsus, with a pair of setae of varying size at the base, or in a
"trident of Neptune" structure, to which the term
"triungulin" refers. The
latral processes, however, have no muscle attachments. The abdominal segments in many species have the tergal and
pleural plates hcavily sclerotized and indurate, whereas the skin of the
intersegmental areas is delicate and white in color. The body consequently has a distinctly
banded appearance after becoming distended with food. In many of the species that attack locust‑egg
masses, this coloration of the segments is not nearly so distinct. One pair
of caudnl cerci is found usually, though not always, on the ninth abdominal
segment, these being very short in some species whereas in others their
length may exceed half that of the abdomen.
In a few species, a supplementary pair is found on the eighth
segment. The various body segments
may bear a number of short setae, usually arranged in transverse rows. The abdomen terminates in a somewhat
bulbous adhesive disk that is utilized in clinging to the substratum. By means of the erectile caudal cerci,
aided by the adhesive disk, the triungulinid is able to assume an upright
position when stimulated by the approach of a carrier, a habit that is
general nmong larvae of this type in several orders. The data available indicate that the number of pairs of spiracles
may be seven or eight, with the first pair occurring on the mesothorax and
the remainder on the successive abdominal segments. In a number of species those of the mesothorax and the first
abdominal segment are very large, and the remainder minute. Several species are known to have the
spiracles of the eighth abdominal segment borne at the tips of a pair of falciform
processes, which are considered to provide against the entry of fluids into
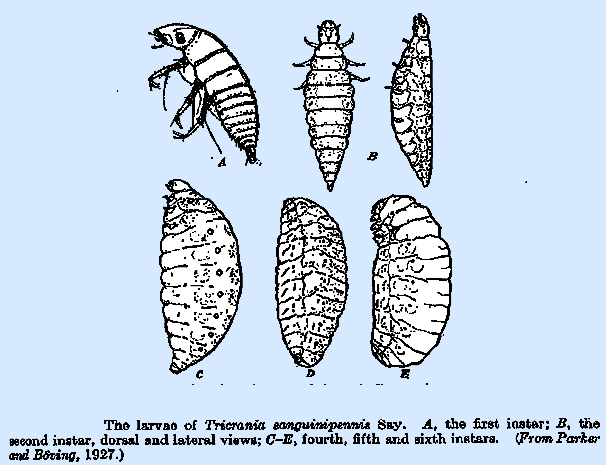
the tracheal system. The second‑instar (caraboid) larva (Fig. 239B) differs from
the preceding instar principally in its more robust body and reduced head and
legs. A rather delicate, white skin
replaces the heavily sclerotized and indurated integument. In Tricrania
(Fig. 24OB) and others that attack bees, the body assume a distinctive boat‑like
form in which the dorsum is nearly flat and the venter markedly convex,
enabling it to float readily upon the surface of the food material in the
cell. This modification in form is
accompanied by a shift in the position of the spiracles, which become
markedly dorsal. The antennae are now
somewhat conical in form and lack the long terminal setae. The integumentary setae may persist,
particularly in the species attacking locust egg-masses, but they are reduced
in size. The third‑ and fourth‑instar (scarabaeoid) larvae
(Fig. 239C) of the species attacking locust eggs are quitc similar in all
respects and have a striking resemblance to the grubs of Scarabaeidae. The abdomen is much increased in size, and
the legs are further reduced. In Tricrania (Fig. 240C), Apalus, and others of the
Nemognathinae, the third instar is quite similar to the second, retaining its
boat like form, but in the fourth instar the abdomen is greatly distended and
markedly convex dorsally as well as ventrally. The fifth‑instar (coarctate) larvae of Epicauta (Fig. 239D) and others of similar host
preferences are markedly different from the preceding instars in having the
legs rudimentary, represented by conical protuberances only, the mouth parts
likewise rudimentary and not serving for feeding purposes, and the body
segmentation often indistinct. A
pronounced longitudinal ridge extends along the lateral margins of thc first
seven abdominal segments. The
integument is much heavier than in the preceding instars and bears transverse
corrugations. Among the species of
Nemognathinae and other groups attacking bees, this instar (Fig. 240D) is not
nearly so distinctly differentiated, though the mouth parts and legs are
greatly reduced and functionless. The
term "coarctate" is not properly applicable to all larvae of the
family of this instar, inasmuch as many are not enveloped by the exuviae. The sixth‑instar (scolytoid) larva is so named because of
its resemblance to the larvae of the Scolytidae, and it thus represents a
reversion to a form somewhat similar to the scarabaeoid. It is of reduced size as compared with the
two preceding instars, owing to the last two transformations having been
accomplished without feeding. The
legs, though still largely functionless, are relatively larger than before. References: Please refer to <biology.ref.htm>, [Additional references may
be found at: MELVYL Library ] |