|
File: <ichne1.ima.htm> [For educational purposes only] Terminology Glossary <Principal Natural
Enemy Groups > <Citations> |
Immature Stages of Ichneumonidae
The immature stages of Ichneumonidae were discussed in detail by
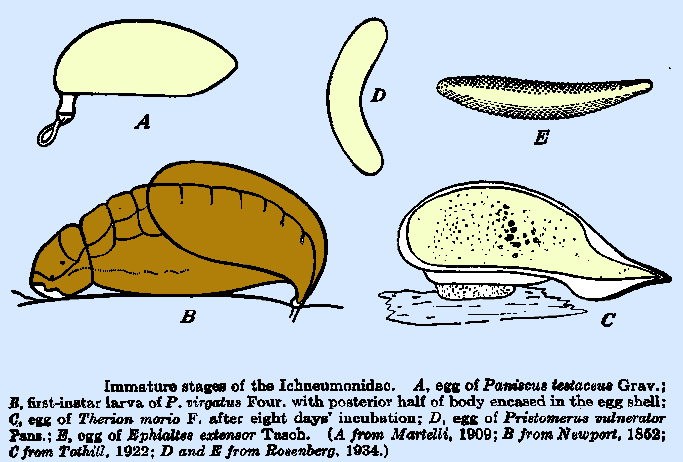
Clausen (1940) as follows: The Egg.-- The eggs of the great majority of species of the
family are of simple form, without a stalk or pedicel and usually with no
sculpturing of the chorion. The shape
is variable, ranging from the broadly oval to cylindrical and, in this simple
form, to the extremely slender forms represented by those of Echthropsis porteri and Perithous mediator Grav., which are only one‑twentieth as wide
as long, curved, and with both ends tapering to points. The eggs of the Cryptinae, Joppinae,
Ichneumoninae, and Ophioninae are, with few exceptions, of the above general‑form. In the latter two subfamilies, the stalked
type of egg is also found, the extreme development of this modification being
in the genera Rhyssa and Megarhyssa, in which the
anterior end is drawn out into a slender tube. The stalk of the egg of R.
persuasoria is approximately
four times the length of the egg body, and the total length of the egg is
12.0 to 13.5 mm. Surface sculpturing on the chorion occurs in only a few species
of the above subfamilies and is not elaborate. In Cryptus sexannulatus Grav. the egg
bears light longitudinal markings, whereas that of Ephialtes extensor
(Fig. 32E) is covered with closely
set "bosses" arranged in rows.
The color is usually translucent white, with the eggs of a few species
assuming a brownish tinge as incubation progresses. Please CLICK on pictures to view details:
The eggs of the Ophioninae are usually of the normal kidney‑shaped
or elongate form, but several genera reveal an adaptation for attaching them
to the integument of the host larva.
This modification is represented by a "pad" or
"button" at the mid‑ventral side of the egg by means of which
it "adheres" to the inner side of the integument of the host at 3
point in the body opposite that at which the ovipositor is inserted. This form is represented by Therion morio (Fig. 32C),
and one that is apparently similarly modified is described by Plotnikov
(1914) in Heteropelma calcator. Please CLICK on pictures to view details:
The most
striking modifications in egg form occur in the ectoparasitic species of the
Tryphoninae and Lysiognathinae; in these groups, the eggs either are partly
embedded in a puncture in the integument of the host larva or have an
adaptive modification of the chorion at the posterior end into some form of
anchor, which is embedded
therein. In the Paniscini, this
adaptation (Fig. 32A) uniformly
appears as a short, blunt pedicel, situated somewhat ventrally, from which
extends a spiral, looped or " braided " process that is stated to
be very elastic at the time of deposition.
Only this latter portion is embedded in the wound. Chewyreuv points out that the pedicel is
not an extension of the egg chorion, for it dissolves completely in potassium
hydroxide. Associated with this form
of egg is a distinctive coloration, the chorion being black or brown and shining,
thus making it conspicuous upon the body of the host. The darkening of the chorion is most
pronounced in the Paniscini and is of varying extent, and at times entirely
lacking, in the Tryphonini. The extreme modifications in egg form are found among the
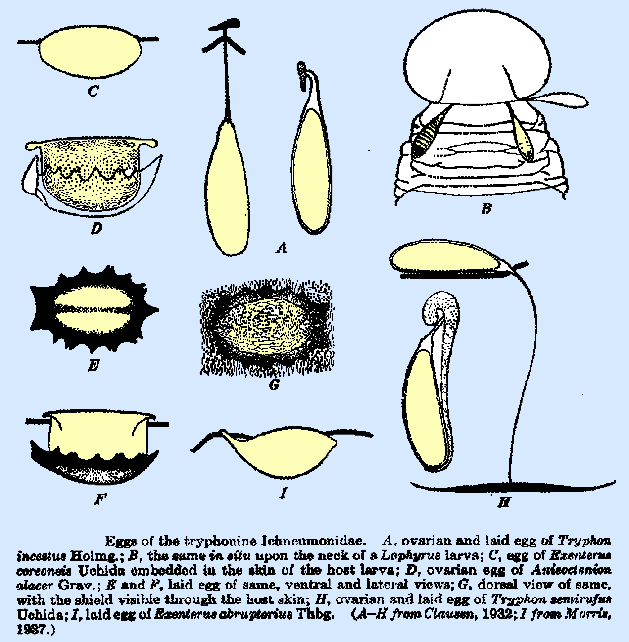
Tryphonini and Cteniscini. Several of these have been described and
figured by Clausen (1932a). The egg
of Tryphon semirufus (Fig. 33H) has a long thread‑like pedicel,
twice the length of the egg body, which bears at its distal end a long,
heavily pigmented bar, attached at the middle and serving as an anchor deep
within the tissues of the host.
Bischoff (1923) figures an identical egg for an undetermined
tryphonine species in Europe; that of T.
rutilator Holmg., the
ovarian form of which is illustrated Pampel, is evidently very much like
it. The egg of T. incestus
(Fig. 33A, B) is of the same general form; but the pedicel is shorter, the
anchor much smaller, and the latter is inserted immediately beneath the
integument. That of Tricamptus apiarius Grav. figured by Bischoff is similar to it. The egg of Exenterus tricolor
Roman (Morris et al., 1937) is of the same general form and bears a scale‑like
sculpturing. In these species, the
chorion is exceedingly heavy and tough and is difficult to puncture, even
with a needle. In Anisoctenion alacer Grav. (Fig. 33D, F), the
anchor assumes a curious and quite different form, in which it is represented
by a blackened shield, with serrate margins, on the ventral side of the egg
body. This shield, which is slightly
larger than the egg, opens out, umbrella‑like, at the time of
deposition. The entire egg except the
dorsal surface lies beneath the host integument, and the exposed portion of
the chorion bears delicate reticulate markings. The egg of Lysiognatha
sp. (Cushman, 1937) is apparently quite similar to that of Tryphon incestus. In all
the species that deposit eggs of the pedicellate type, the adaptations that
will appear in the laid egg can be detected by an examination of the ovarian
egg (Fig. 33D, H). Not all the Tryphoninae possess eggs of the pedicellate type
discussed above. In the Diplazonini,
the egg is ellipsoidal in form, with both ends smoothly rounded. That of Hypamblys albopictus
is kidney‑shaped, whereas the egg of Exenterus coreensis
Uchida (Fig. 33C) is oval in outline,
with no pedicel whatever, and is largely embedded in the wound. The egg of E. abruptorius
(Fig. 33I) described and figured by Morris (1937) may be considered as
intermediate in form between that of E.
coreensis and of Tryphon incestus, and it shows an incipient pedicel
formation. This is represented by a
slender cylindrical extension at the posterior end. At oviposition, the body of the egg is largely embedded in the
wound, only a portion of the dorsum being exposed through the aperture in the
skin, and the tip of the pedicel also protrudes, though from a separate and
minute hole. The variation in egg
form and manner of deposition within a genus is illustrated by the three
species of Exenterus that
have been mentioned. First‑instar Larva. --What may be termed the normal
hymenopteriform first instar larva of the family is that of the ectophagous
species of the Ichneumoninae and other subfamilies; it is characterized by a
large and often heavily sclerotized head, with large conical antennae and
simple mandibles, and 13 body segments of diminishing width. The integument may be bare or clothed with
numerous minute spines. Several species
that develop internally are of this same general form. The first‑instar larva of P. nigridens bears six pairs of small setae on each segment;
in addition, each abdominal segment bears a broad transverse band of minute
integumentary setae. The anal opening
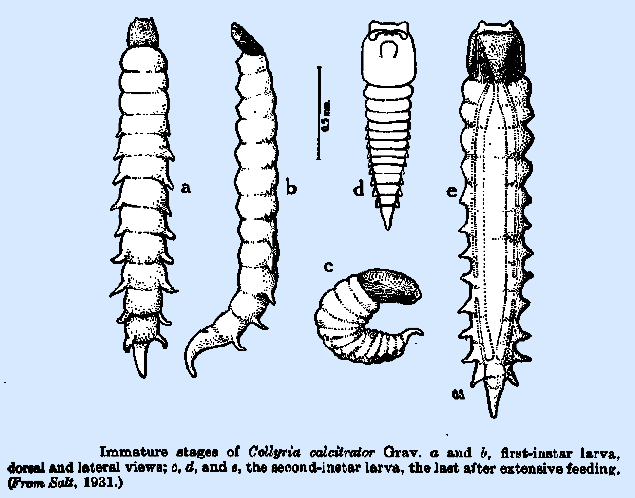
is usually situated dorsally, though it is said to be on the venter of the
thirteenth segment in C. calcitrator (Fig.
34a). In this
species, paired fleshy processes occur dorsolaterally on the abdomen; they
are of increasing length on the successive segments. In the Paniscini, the first‑instar larva has been described
only for Paniscus cristatus Thoms. It differs from the normal hymenopteriform
larva only in the possession of numerous forward‑directed spines on the
venter and sides of the last abdominal segment, an adaptation to hold the
caudal end of the body more firmly within the eggshell during
development. Polysphincta, which has the same habit, is not known to
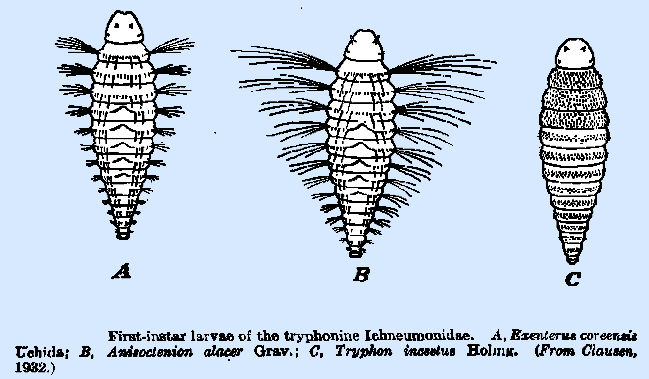
possess this character. The first‑instar larva of Anisoctenion alacer
(Fig. 35B) is markedly different from those thus far discussed, though still
of the hymenopteriform type. Each
body segment bears a transverse row of long hairs at each lateral margin;
these are of decreasing length and number on the successive segments. Each of the first five abdominal segments
bears a pronounced welt on the median dorsal line. This larva normally moves upon its back in a looping manner,
the welts and the caudal sucker aiding in accomplishing locomotion, while the
lateral tufts of long hairs hold the body in a horizontal position. Exenierus coreensis (Fig.
35A) and several others of that genus and Tryphon semirufus
have similar larvae, though the lateral tufts of hairs on the latter are much
shorter. The larva of Tryphon incestus (Fig. 35C), however, lacks both the dorsal welts
and the lateral tufts of long hairs, is densely clothed with minute spines,
and does not assume an inverted position when in movement. The most common type of first‑instar larva among the
endoparasitic species is the caudate, which attains its highest development
in Ichneumonidae. The body is
somewhat cylindrical, with 11 to 13 recognizable segments, and the integument
is usually smooth and shining. The
tail may equal or exceed the body length, and it may be slender and taper to
a sharp point or be almost cylindrical, with the distal end broadly rounded,
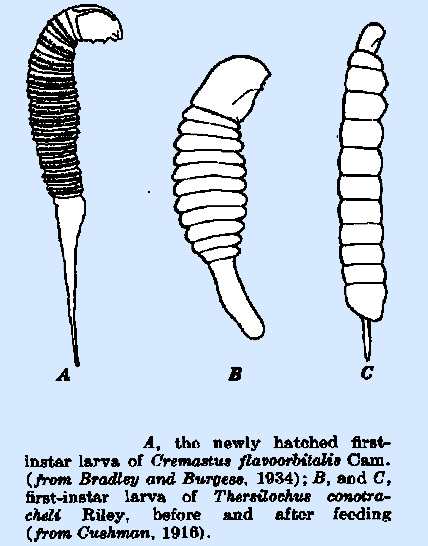
as in Thersilochus conotracheli (Fig. 36B,
C). In some species, as Anomalon cerinops Grav, the terminal portion of the tail is
spined. Timberlake (1912) considered
the tail of Eulimneria valida Cress. to be a blood
gill, whereas the extensive ramifications of the tracheal branches in the
tail, illustrated by Tothill (1922) in the larva of Hyposoter pilosulus,
which led him to attribute a respiratory function to that organ, have been
shown by Thompson and Parker to represent an erroneous interpretation of the
structures observed in mounted specimens.
Working with Eulimneria
crassifemur Thoms., a
species of very similar form, they determined that the supposed bundle of
tracheids is simply a lobe of the fat body from which the fat globules have
been dissolved by the reagents employed. Thorpe (1932) has studied the tail appendage of a series of
species of this and other families with particular reference to its role in
respiration. He found an appreciable
variation in the extent to which the tracheal branches extend into this
organ. In the majority of species, the
lateral tracheal trunks extend into it and terminate in the fat body, but in Cremastus interruptor they branch and extend through the basal two‑thirds
of the tail. The newly hatched caudate larvae of Cremastus flavoorbitalis
Cam. (Bradley and Burgess, 1934) (Fig. 36A) and C. interruptor
Grav. bear a double row of scallops transversely on each body segment; these
disappear before the first molt and are believed to be an adaptation to
permit of rapid increase in body size.
The larva of Anomalon
cerinops has a pair of small
slender processes ventrally on the first and third thoracic and the sixth and
eighth abdominal segments. The first‑instar larva of Omorgus mutabilis
Holmg. bears a pair of prominent tusk‑like sense organs on the head
that project downward and backward from the posterior ventral margin of the
head capsule. They represent one of
the four pairs of sense organs present on the venter of the head of larvae of
this family. Many of the caudate larvae have the head comparatively large,
heavily sclerotized, with falcate mandibles, approaching that of the
mandibulate type. The larva of Syrphoctonus maculifrons Cress. may properly
be considered as of the latter type, for the head is equal to the thoracic
region in width and the tail is hardly evident (Kamal, 1939). It bears a strong resemblance to the
mandibulate larvae of the Braconidae, particularly of Opius. In Diplazon and Homotropus, of the same
subfamily, the head is smaller and the tail more fully developed, though
still short. The vesiculate type of larva is not nearly so common, nor is the
vesicle so highly developed as in the Braconidae. Usually it is in an incipient stage, is small in size, and
often is not readily recognized because of being retracted at the time of
examination. A number of the caudate
larvae of the Ichneumoninae and Ophioninae, such as Glypta rufiscutellaris,
Nemeritis canescens, and Anomalon cerinops, bear the vesicle dorsally at the base of the
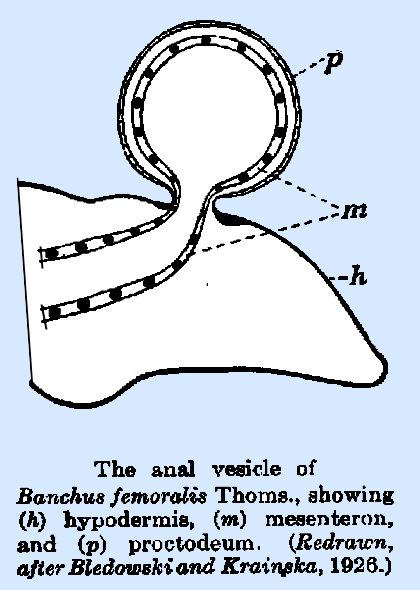
tail. A typical ichneumonid vesicle
is that of Banchus femoralis Thoms., illustrated
in Fig. 37. The polypodeiform type of larva is found in Hypamblys albopictus
(Wardle, 1914) in which the paired thoracic processes are lobe‑like and
those of the abdominal segments rather sharply pointed. The tail is approximately one‑fifth
the length of the body. There is apparently no essential distinction between the
respiratory systems of ecto‑ and endoparasitic first‑instar
larvae. Some are stated to be
entirely devoid of tracheae~ whereas others have a complete internal system
corresponding to that of the mature larva except for the lack of
spiracles. The tracheal system of Phaeogenes nigridens, which has been fully described by Smith,
consists of a main lateral trunk on each side of the body connected by main
transverse commissures dorsally in the first thoracic and ventrally in the
ninth abdominal segment. Accessory
lateral commissures connected with the main trunks by three branches, extend
from the posterior margin of the first thoracic to the anterior margin of the
first abdominal segment. In each of
the first nine abdominal segments, the ventral branches are connected to form
secondary transverse Gommissures. With very few exceptions, the first‑instar larvae of this
family lack spiracles. Paniscus cristatus is said to have a pair on the prothorax; Meyer
(1922) illustrates that pair, and eight additional pairs on the abdomen, in Tryphon signator Grav. Imms (1918b) found nine pairs of spiracles
on the first‑instar larva of Pimpla
pomorum; Speyer (1926),
studying the same species, noted an additional pair, very minute, on the
thorax. The general lack of an open
tracheal system is in contrast to the Braconidae and other extensively
studied families of the order, in which the ectoparasitic first‑instar
larvae are quite consistently provided with open spiracles. Intermediate‑instar Larvae. --The information available as
of 1940 was insufficient to make an adequate comparison of the larval
instars between the first and last, due primarily to uncertainty as to the
total number. A considerable number
of species are stated to have only three instars, and others four; many are
known to have five instars.
Unquestionably, some of those said to have only three will reveal, on
closer examination, a greater number.
Rosenberg mentions the occurrence of six instars in occasional larvae
of Cryptus sexannulatus Grav. and Hemiteles hemipterus, though the normal number is five and four,
respectively. In the species of
Paniscini and Polysphinctini that retain connection with the eggshell during larval
development, the number of instars can be readily determined by a count of
the exuviae forming the pad beneath the posterior portion of the body. In species having hymenopteriform first‑instar larvae,
there is little change in general form in the following instars, but those of
caudate form in the first instar usually show a progressive reduction in the
appendage, with its complete absence in the last instar. In Thersilochus
conotracheli, it disappears
entirely with the first molt, and in some other species it persists only
through the second instar. The
bidentate mandibles appear in the second instar in Ephialtes examinator. The second‑instar larva of Collyria calcitrator (Fig. 34c, e) is of a pronounced mandibulate
type, with the head wider than the body and the mandibles large and falcate
in form. The fleshy dorsolateral
processes on the abdomen persist in this instar. The stage of development at which the spiracles appear is
variable. In Ephialtes examinator
and Phaeogenes nigridens the nine pairs are
evident in the second instar, though in the latter species, which is
internal, they are nonfunctional. Angitia fenestralis Holmg. reveals the spiracles in the
penultimate instar, but in the majority of species they appear only in the
last one. Mature Larvae. --The normal last‑instar larva of the
Ichneumonidae has 13 distinct body segments, the integument usually smooth
and glistening, and it bears no fleshy processes or appendages. In Phaeogenes
nigridens, there is a very
characteristic dorsal hump on the third thoracic and first abdominal
segments, a modification in form said by Smith to be necessary because of the
manner of feeding of the larva. In
the majority of species, the mandibles are simple, often with minute spines
on the margin, though a few are bidentate and those of Echthropsis porteri
are 5-dentate. In Xylonomus brachylabris Kr., the mandible has a concavity on the
inner side flanked by ridges crowned with distinct teeth. The mandibles of Polysphincta are stated to be curved outward at the tips,
and the puncture in the host integument is made, not by a pinching action,
but by the tips being brought together, inserted, and then spread apart. Each body segment usually bears a row of
small, delicate spines transversely that may encircle the segment. In Philopsyche
abdominalis Morley (Skaife
1921b), there are two distinct bands of short spines on each segment, those
of the first band being directed cephalad and those of the posterior band
caudad. This is presumably an
adaptation for movement within the case of the bagworm host. The larva of Pimpla pomorum
bears numerous minute papillae upon the integument. The tracheal system consists of the two main longitudinal trunks
connected by dorsal anterior and ventral posterior commissures, with a
supplementary lateral trunk on each side extending from the posterior margin
of the first thoracic segment to the anterior margin of the first abdominal
segment and connected with the main trunk by three branches. There are usually nine pairs of spiracles,
the first of which, though mesothoracic in origin, is situated at the
posterior margin of the prothorax, the remainder being near the anterior
margin of the first eight abdominal segments (Fig. 38). Angitia
fenestralis (Meyer 1915) is
stated to have 11 pairs of spiracles, situated on all thoracic and the first
eight abdominal segments. Imms
(1918b) called attention to the occurrence of 10 pairs in Pimpla pomorum, the additional pair on the 2nd thoracic segment
being vestigial and nonfunctional.
Thorpe (1930) mentioned this in a discussion of P. ruficollis
Grav. and stated that the occurrence of the vestigial pair on the 2nd
thoracic segment is probably general in the family but has been largely
overlooked. There are 10 pairs in Polysphincta tuberosa, also, but those of
the thorax are on the 1st and 3rd segments, while in Collyria calcitrator
(Fig. 38) they occur on the 2nd and 3rd.
The tracheal system of the latter species differs also from the normal
for the family in the lack of the lateral accessory and the posterior ventral
commissures. Salt pointed out the
general similarity of the larval characters of the species to those of the
Braconidae. In Scambus detrita
and other species, the ventral branches in each abdominal segment unite to
form accessory ventral commissures. The greatest modification in mature larval form and in functional
adaptation occurs in the tribe Polysphinctini and in certain other
Ichneumoninae. These species are
parasitic upon spiders or are predaceous in their egg capsules. The morphological modifications are of two
forms and serve distinct purposes.
The first of these is the occurrence dorsally of retractile
"welts" (Fig. 39C), surmounted by a number of hooked spines or of
patches of straight spines, which serve to hold the larva in the web during
the spinning of the cocoon or to facilitate movement in the egg capsule. The second modification is the development
of paired fleshy processes ventrally on certain abdominal segments to attach
the body firmly to the exuviae and thus to the body of the host spider. The mature larvae of a considerable number of species have been
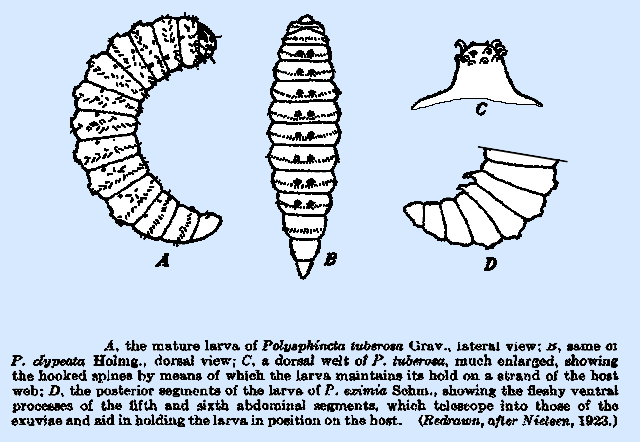
described by Nielsen (1923), and the dorsal welts, bearing the hooked spines,
occur in most if not all species of Polysphincta,
Schizopyga, and Zaglyptus. The number of welts is usually seven or
eight, and they occur in a single row on the median line of the third
thoracic and the following seven segments in P. tuberosa
Grav. (Fig. 39A), P. eximia Schm., and P. nielseni
Roman. Four welts only are recorded
on the larva of P. gracilis Holmg., whereas in P. clypeata Holmg.
(Fig. 39B), P. pallipes, and S. podagrica Grav. they are paired, rather than single, on
each segment. In the last species,
they occur on the first six abdominal segments (Nielsen, 1935). Laboulbene (1858) records them on the
first seven body segments in P.
fairmairii Lab., and Maneval
(1936) stated that they are on the first seven abdominal segments in Z. variipes Grav. In
these two species, also, the welts are single rather than paired. The hooked spines that surmount each welt
are directed outward from the center of the welt; and when one of these, or
more, is drawn over a strand of the host web and the welt then retracted into
the body, the larva is very securely held in position. In Tromatobia
oculatoria F., the spines
are simple and straight and arranged in transverse bands at the anterior and
posterior sides of the welt. Those at
the front are directed cephalad, and those at the rear caudad. Many if not all of the species of Polysphincta have a pair of fleshy conical processes (Fig.
39D) ventrally on the fifth and sixth abdominal segments, and these are
embedded in the exuviae beneath the body.
They are present upon the intermediate instars, also. In S.
podagrica, there are four
pairs of these processes rather than two, and they occur on the fifth to the
eighth abdominal segments. References:
Please refer to <biology.ref.htm>,
[Additional
references may be found at: MELVYL Library ] |