|
File: <encyr1.ima.htm> [For educational purposes only] Terminology Glossary <Principal Natural
Enemy Groups > <Citations> |
Immature Stages of Encyrtidae
Immature
stages of Encyrtidae were discussed in detail by Clausen (1940), as follows: The Encyrtidae reveal an
exceptional diversity in form of the immature stages, and many of the
modifications are strictly adaptive.
These are made necessary not only by the wide range of hosts attacked,
but by the varied conditions under which development takes place. The Egg. --Two general types of
egg are produced by the Encyrtidae, there being the stalked and the
encyrtiform, the latter representing an adaptive modification of the
first. In both forms, the ovarian egg
is two‑bodied, and the contents of the anterior body, or bulb, are
forced into the egg proper at the time of oviposition, leaving the stalk as a
slender tube at the anterior end. In
the stalked form, this stalk is functionless after deposition but may, in
some instances, serve to attach the egg to the integument or to other of the
internal organs of the host.
Representative genera having this type of egg are Aphidencyrtus, Cerapterocerus,
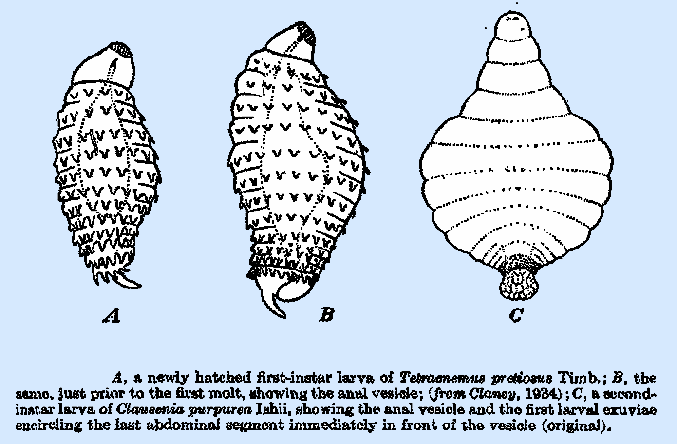
Eusemion, and Anarhopus. In Tetracnemus
pretiosus, according to
Clancy, the stalk is reduced to a broad, blunt petiole one‑third to one‑fourth
the length of the egg body. Please CLICK on
pictures to view details:
Please CLICK on
pictures to view details:
The
encyrtiform egg is distinguished from the stalked form by a heavy surface
rib, termed the aeroscopic plate by Silvestri (1919), which extends the
length of the stalk and of the greater portion of the egg itself. Well‑known genera having encyrtiform
eggs are Encyrtus, Microterys (Fig. 73A, B), Aphycus, Metaphycus, Blastothrix,
and Ooencyrtus. The plate of O. johnsoni
is described by Maple as granulate in appearance and is composed of a mosaic
of cells upon the outer surface of the stalk and egg body except for a
thickened area near the base of what remains of the bulb of the ovarian
egg. In deposited eggs, the plate is
much darker than the remainder of the chorion. In two species of Isodromus parasitic in Chrysopa larvae, it has been
found by Clancy that the egg of one, I.
niger Ashm., is typically
encyrtiform, whereas the other, I.
iceryae How. (Fig. 74),
lacks the aeroscopic plate and bears merely a melanized ring and a delicate
membranous collar on the stalk. First‑instar Larvae. --Among
the monoembryonic species of the family, four forms of first‑instar
larvae may be distinguished, based upon morphological modifications having a
functional nature. The
hymenopteriform larva has a body of 12-13 visible segments, is widest in the
thoracic or anterior abdominal region, and has no sculpturing or segmental
processes. These larvae lie free in
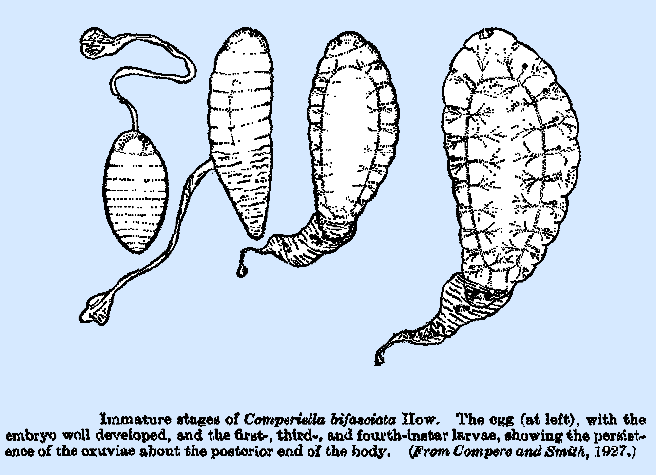
the body cavity of the host and lack the open tracheal system. A typical representative of this group is Comperiella bifasciata How. (Compere and Smith, 1927). The second is the encyrtiform, so
called because it hatches from the encyrtiform egg previously described. The number of body segments is reduced,
there being only 10-11 visible, and the last segment, which bears the single
pair of spiracles, apparently represents several that have fused. The last four or five segments are usually
closely enveloped by the eggshell, and this connection persists through the
greater portion of the larval stage.
The larva of Isodromus
iceryae, which hatches from
the modified encyrtiform egg already described, is hymenopteriform, for it
lacks the posterior spiracles and consequently does not derive its air supply
through the stalk. Both the egg and
first‑instar larva of this species appear to represent transitional
stages between the hymenopteriform and the encyrtiform type. The genus Microterys contains many well‑known species having
encyrtiform larvae; yet it has been shown by De Bach that in M. titiani it is hymenopteriform, with a full complement of
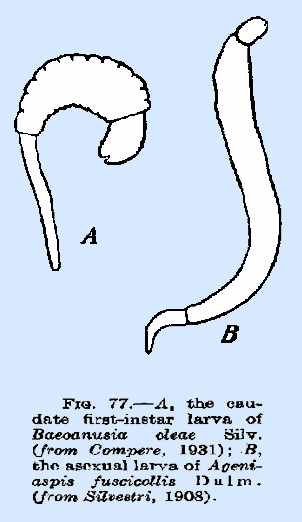
spiracles. The caudate larva (Fig. 77A) is
frequently found among the species attacking Coccidae and Aphididae and is
characterized by the development of the last abdominal segment into a tail‑like
organ that may exceed the body proper in length and may bear setae on the
distal portion. It is associated
with the stalked type of egg. These
larvae do not possess open spiracles.
Many genera have larvae of this type, the best known being Aphidencyrtus, Cerapterocerus, Cheiloneurus, Eusemion, and others. The vesiculate form is similar to
the hymenopteriform, except that the proctodaeum is evaginated to form a
caudal vesicle. This modification is
rare among the Encyrtidae and is at present known only in the genera Anarhopus and Clausenia (Fig. 76C), both of
which parasitize mealybugs. Tetracnemus pretiosus (Fig. 76A, B) may be
of the same type, though it is uncertain whether or not the small expanded
organ on the caudal segment corresponds to the vesicle in the above‑named
species. Both A. sydneyensis
and T. pretiosus
are distinguished from other known Encyrtidae by the presence of a ring of
fleshy processes or protuberances on the first 12 body segments; and the
former has also a single, curved medium process dorsally on the last segment,
immediately above the vesicle. Intermediate‑instar and
Mature Larvae. --The greatest diversification in form occurs in the first
instar, and the succeeding forms tend to become more uniform as the final
instar is reached. The
hymenopteriform larva, which lies free in the body cavity of the host,
progresses through the series of molts without appreciable change in its
essential characters. In the caudate
forms, the tail becomes considerably reduced in size in the second instar and
practically disappears in the third.
The vesiculate forms, on the other hand, show an enlargement of the
vesicle in the second and third instars.
In Anarhopus and Tetracnemus, the ring of fleshy
protuberances on each body segment of the first‑instar larva is lacking
after the first molt. The number and position of the
spiracles of the larvae are an exceedingly variable character in
Encyrtidae. In the hymenopteriform
larva, the spiracles are lacking in the first and second instars, but they
appear on the second to the tenth body segments in the third or a later
instar. Among the species having
caudate larvae, Cerapterocerus mirabilis
Westw. is stated to lack spiracles until the fourth instar, at which time the
nine pairs appear in the position already mentioned. In Carabunia
myersi (Fig. 73), they are
first found on what is stated to be the third and final instar, and only
three in number, the anterior pair being on one of the thoracic segments and the
remaining two pairs on the abdominal segments immediately preceding the
caudal appendage Information regarding the spiracle
arrangement of vesiculate larvae is available only for Anarhopus sydneyensis. In this species, they are lacking on the
first instar and occur on the second to tenth body segments of the second and
last instar. Tetracncmus pretiosus
has no open spiracles until the final instar, when the full complement
appears. The first‑instar encyrtiform
larvae possess a single pair of spiracles on the last apparent abdominal
segment. This arrangement persists in
the following two instars, and the nine pairs of spiracles then appear on the
fourth instar. In Microterys speciosus (Ishii, 1923), they are stated to appear on the
third instar. Clancy mentioned that
the second instar of Isodromus
is readily distinguished from the first by the presence of the spiracular
spurs in the second to ninth body segments, and this character may be common
to many second‑instar larvae of the encyrtiform type. A marked departure from the normal for the
family occurs in Metaphycus lounsburyi (Smith and Compere,
1920), in which the single caudal pair of spiracles of the first instar is
followed by three additional pairs, situated on the second to fourth body
segments, on the second instar, and by the usual nine on the third
instar. A further modification in
spiracular arrangement is found in certain species of Encyrtus having encyrtiform larvae, which acquire in their
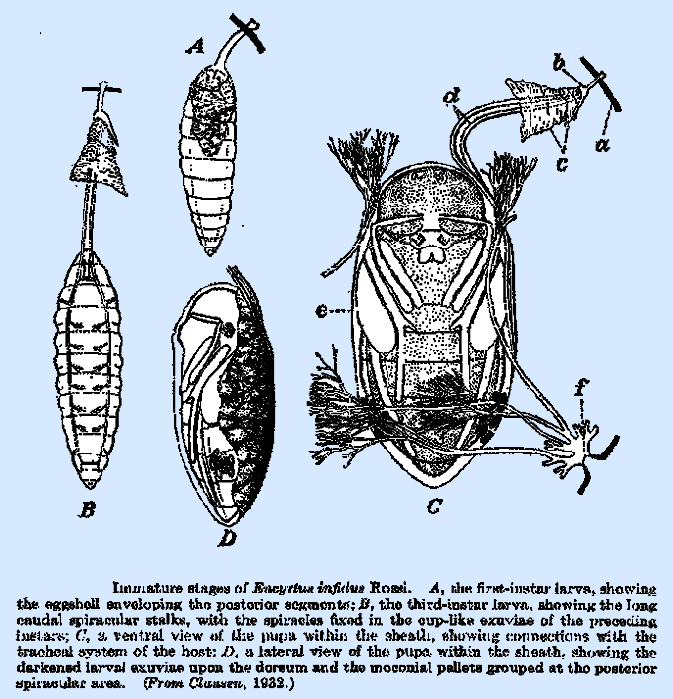
later stages an intimate connection with the host respiratory system. In E.
infelix (Thorpe, 1936), the
fourth instar has 3 pairs of spiracles, one of which is on the prothorax and
the remaining two at the posterior end of the abdomen. The caudal spiracles are borne at the end
of a pair of slender tube‑like processes, merely enclosing the tracheal
tubes, half to‑two‑thirds the length of the body proper. The fifth‑instar larva bears only
two pairs of spiracles, one at each end of the body. This spiracular modification occurs also
in E. infidus, though one instar apparently was overlooked, and
the described third (Fig. 70B) is identical with the fourth of E. infelix. Ishii
(1932a) described the supposed first‑instar larva of E. barbatus Timb. which has the three pairs of spiracles
arranged in identically the same manner as is given above for the fourth
instar. The large size of this larva
indicates that it may be a later instar than that stated (Clausen 1940). In considering the various
adaptations, it is seen that the characters mentioned are common to a number
of genera and that, in some instances, all species of a given genus do not
reveal the same modifications. References:
Please refer to <biology.ref.htm>,
[Additional
references may be found at: MELVYL Library ] |