|
File: <cyrti1.ima.htm> [For educational purposes only] Terminology Glossary <Principal Natural
Enemy Groups > <Citations> |
Immature
Stages of Cyrtidae
First instar larvae of Cyrtidae
are of the planidium type, and are able to undergo a free living period of
considerable duration without food.
Locomotion is either by a looping motion or by jumping. In the latter case, the larva of Pterodontia stands erect on the
caudal disk, which is specialized for the obvious purpose of giving a firm
hold on the substrate, and projects itself into space by the sudden downward
thrust of the long caudal cerci, bringing them into line with the
longitudinal axis of the body from a position at right angles with it
(Clausen 1940/62). However, Bovey
(1936) found this to be accomplished in O.
pallipes by the lowering of
the head to the substrate near the point of attachment of the caudal disk,
thus forming almost a circle, following which the body was abruptly
straightened. This action simulates
that of a released spring. The
planidia of Opsebius are
able to crawl in a looping manner along a single strand of a spider web (Cole
1919). Jumping does not seem induced
by the host presence, but it serves to accomplish dispersion. Actual host contact is mostly by chance,
and the planidia, when alert and awaiting a host or stimulated by its
presence in the immediate vicinity, assume an erect position (Clausen
1940/62). It is not know exactly how long
the planidia are able to life if contact with a host is not made. One weeks seems to be the maximum, which
is short in comparison with that for planidium larvae of other groups of insects. The planidia enter the bodies of their
hosts probably through the thin membranes of the articulations of the
legs. While doing so they cause
appreciable irritation to the host spiders, and Theridium adults bearing planidia of Opsebius scratched themselves excitedly, seemingly being
able to kill some of the parasitoids with their mandibles (Cole 1919). Please CLICK on
picture to view details:
King (1916) recorded finding the
planidia of Pterodontia in
all parts of the host body, even in the legs and palpi, although the majority
were in the thorax. They persist in
this stage, without much growth or direct relation to any host organ, until
springtime. Millot (1938) found Ogcodes pallipes young larvae in the abdomen during winter, but
they migrate to the respiratory organs in the early spring and the first molt
follows. Part of the second stage is
also passed in the abdomen, and the second molt occurs there. The larva of Astomella lindenii
Er., while in the host abdomen, derives air from one of the lung chambers of
the host by placing the caudal spiracles against the lung wall and seemingly
making a minute perforation (Brauer 1869a).
This also occurs in O.
pallipes, although Millot
(1938) found some individuals to perforate the abdominal wall instead and
thereby make direct connection with the outside air. In the latter, there is evidence of the
formation of a respiratory funnel which is comparable to many Tachinidae. Third instar larvae complete their
feeding quickly and consume the entire contents of the host abdomen. Emergence is through a hole cut in the
ventral abdominal wall. The host
spider usually shows no evidence of parasitism, either by modification in
body form or a change in activities, until just a few hours before it
dies. However, the spider does spin a
mat of silk just before death, and Locket (1930) noted that this was done
even by male spiders, which was abnormal.
He attributed this action in both sexes to increased pressure within
the abdomen from the large parasitoid body; a measure of relief is probably
obtained by expelling the silk. Pupation.-- Pupation is outside
the host body, in either the web or the burrow or in some sheltered place
near the place of host demise.
Emerged larvae of Acrocera
fasciata Wied. suspended
themselves in the host web by clinging to a strand with their mandibles; they
were supported also by strands beneath the body (Emerton 1890). A sticky body surface aids the mature
larvae to retain their positions in the web or on a trunk. Actual pupation usually occurs the day
following emergence from the host, and the meconium is cast by the
prepupa. The pupal stage is short,
being 7 days in Pterodontia,
6-11 days in O. pallipes, and about the same in
several other species in summer.
Adult emergence occurs through a median longitudinal break over the
prothorax of the pupal shell. There
seems to be only a single generation annually, 9 or more months being passed
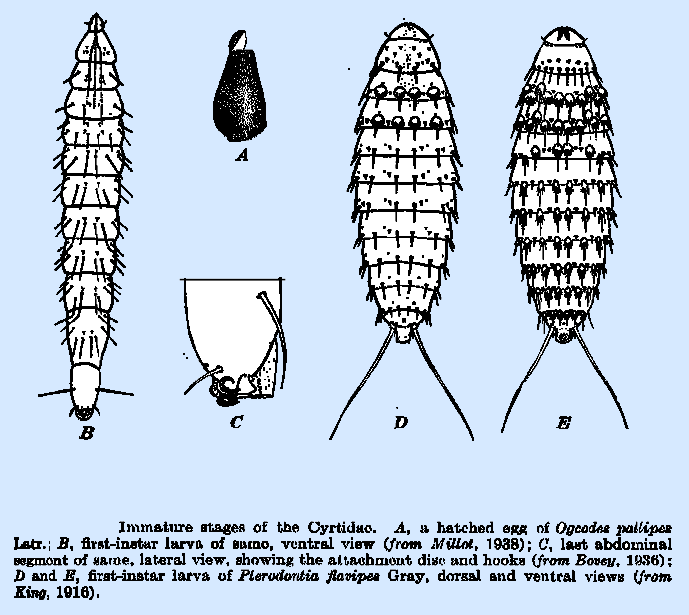
in the first larval stage within the active or hibernating spider. The eggs of all species of
Cyrtidae thus far described are of minute size, rarely exceeding 0.25 mm. in
length, pear‑shaped in outline, and brown to dull black in color. Millot called attention to an adhesive
disk at the larger, posterior end of the egg of O. pallipes (Fig. 162A) by means
of which it becomes firmly attached to any object upon which it may be
deposited. The first‑instar larvae have
been described for O. pallipes Bovey 1936; Millot
1938), Ogcodes sp. (gibbosus L. or zonatus Er.; Konig 1894), and P. flavipes. All are
distinctly of the planidium type, are 0.3 to 0.4 mm. in length, and have
heavily sclerotized segmental plates both dorsal1y and ventrally. In Pterodontia,
the body is fusiform, whereas in others it is more elongated, approaching the
vermiform. P. flavipes
(Fig. 162D, E) is distinguished not only by the exceptiona11y long caudal
setae, or stylets, nearly half the length of the body, which in other species
do not exceed the length of the last abdominal segment, but by the palmate
scales which occur in transverse rows upon the dorsum of the thorax and
ventrally on all body segments except the first and last. In other described species, these scales
are replaced by simple though heavy spines.
0. pallipes (Fig. 162B) and
several others of that genus have two pairs of spines, rather than one, on
the caudal segment. The head is relatively small, and,
in 0. pallipes, the mouth hook terminates in a sharp, upwardly
curved point and the three pharyngeal plates are long and slender. The last abdominal segment of 0. pallipes (Fig. 162C) is highly developed as an attachment
organ, consisting of the usual adhesive disk and, in addition, three strong
hooks, of which the outer two are simple and the median one bifurcate. These structures apparently relate to the
manner of jumping and serve to brace and hold the body more firmly than is
possible with the disk alone. There is a difference of opinion
regarding the occurrence of an open respiratory system in the first‑instar
larvae of this family. Konig stated
that Ogcodes sp. bears a
single pair of spiracles in two funnel‑shaped depressions at the
posterior margin of a triangular "shield" on the penultimate
abdominal segment. In 0. pallipes, they are situated dorsolaterally upon the
anterior portion of the last segment.
King, however, noted that spiracles are lacking in Pterodontia and that the two
crescentic areas at the posterior margin of the eighth abdominal segment,
which superficially resemble spiracular openings, are in reality only the
notches in the dorsal plate in which the caudal cerci fit when the larva is
in the erect position. The second‑instar larva was
not described for any species when Clausen (1940) published his book, though
several authors had stated that it has a very close resemblance to the third
instar. The fully fed third‑instar
larvae of Ogcodes costatus (Fig. 163) and P. flavipes are rather robust, rather indistinctly segmented,
and widest in the mid‑abdominal region and have a marked constriction
between the thorax and abdomen (Kaston 1937). The posterior portion of the abdomen tapers sharply, and the
last segment is broadly conical. The
integument is delicate and entirely unornamented except in 0. pallipes, which has two longitudinal bands of minute
"granulations" along each side of the abdomen, and P. flavipes, which has pairs of pad‑like areas armed
with crochets on the ventral surface of the 5th to 11th body segment. The mouthparts are small and relatively
simple. In 0. costatus and O.
pallipes, spiracles are
present on the first thoracic and the eighth abdominal segments only, whereas
the anterior pair of Pterodontia
is believed to be on the mesothorax.
This pair of spiracles is apparently not functional in 0. pallipes and Pterodontia. There is a marked similarity in
form between the pupa of all species that have been studied. The variable characters noted are the
crest, or groups of presumed sensory organs on the head, and the number of
spiracles. The V‑shaped crest
of Pterodontia consists of a
large number of club‑shaped processes; in 0. costatus (Fig. 163) and 0. pallipes, they occur in two groups on each side of the
head. Astomella lindenii
lacks the crest, or groups of processes on the head, and has a row of spines
on the dorsomeson of the thorax.
Brauer recorded six pairs of abdominal spiracles in Astomella, several species are
thought to have four pairs, and King mentioned three in Pterodontia.
Millot found four functional pairs on the anterior abdominal segments
of 0. pallipes, followed
by two vestigial pairs on the next segments.
He called attention to the occurrence of only four pairs of abdominal
spiracles on the adults of this family and consequently consider it probable
that the pupae will be found to have a like number. References:
Please refer to <biology.ref.htm>,
[Additional
references may be found at: MELVYL Library ] |