File:
<zygomycota.htm> <Index to Mycology> Pooled References <Glossary> Site
Description <Navigate to
Home>
Page 2
True Fungi (Eumycophyta) 1
Zygomycota (Phycomycetes) -- Zygote fungi
Biflagellatae:
(Contact) Sample
Examinations
CLICK on illustrations to enlarge: Biflagellatae The principal orders in the Biflagellatae are Lagenidiales, Saprolegniales, Leptomitales and Peronosporales, in a presumed
increasing order of evolution. This is
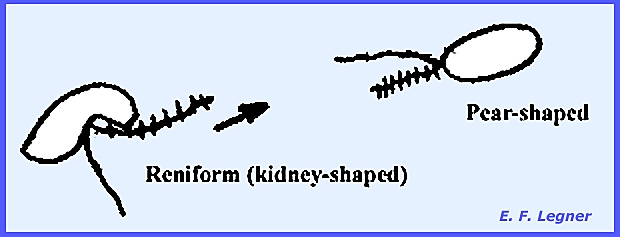
the last group in the fungi whtat possess flagellated cells at all. Spores may be either of two kinds: pear shaped or reniform (kidney-shaped). When a tinsel flagellum is present it is
always to the anterior portion of the spore.
Flagellated cells in the Biflagellatae are always zoospores, and there
are no flagellated gametes.
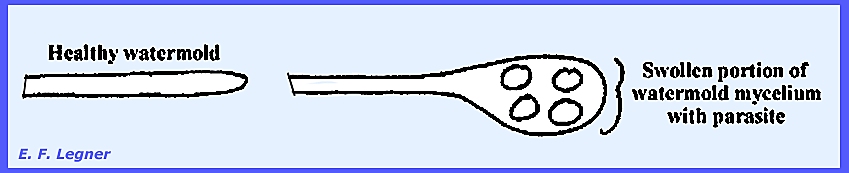
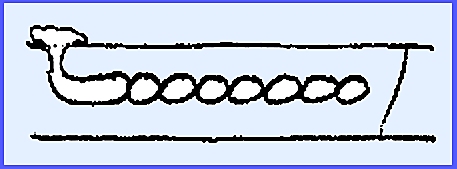
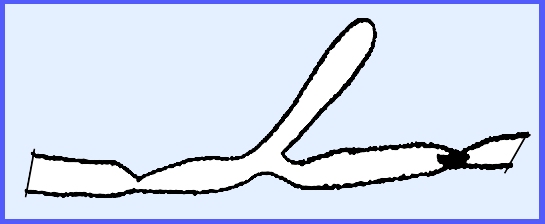
In the order Lagenidiales Olpidiopsis vexans is
a typical representative species. It
is a simple, holocarpic, biflagellated form with no mycelium. It parasitizes water molds causing a
hypertrophy of the mycelium. Two
principal families are Olpidiopsidaceae and Lagenidiaceae.
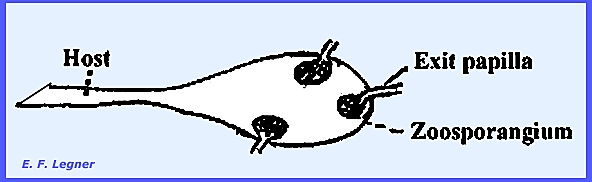
The entire thallus forms
zoosporangia.
Zoospores escape and reinfest a
water mold
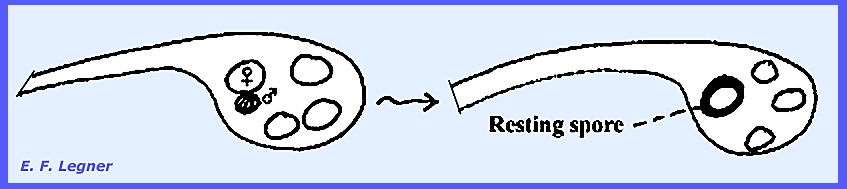
The gametangia are differentiated
into male and female and copulation occurs, the contents of the male empties
into the female.
Resting spores are released after
decompositon of the host and later zoospores are produced. ---------- Another example of Lagenidiales is
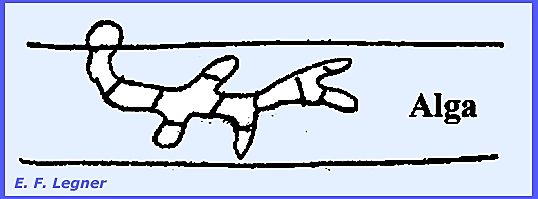
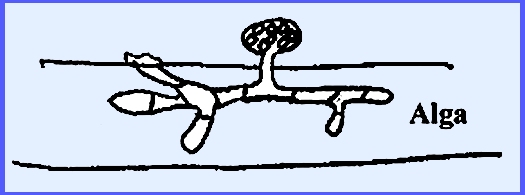
Myzocytium
proliferum. This
species parasitizes algae of the genus Spirogyra. A zoospore lands on a filament, germinates into a mycelium,
which then septates.
The chloroplast or spiral of the
alga collapses after infection. The
segments each round off and form gametangia.
This is a holocarpic organism.
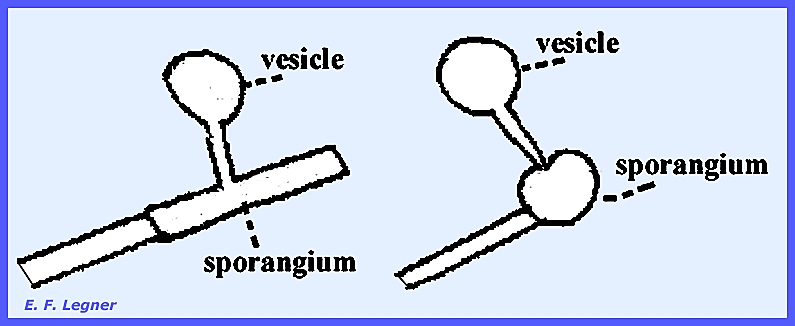
An exit papilla is formed on each sporangium
and zoospores fill into a balloon-like vesicle in the immature stage. Later on maturing the vesicle bursts and
the zoospores are scattered.
Any two of the sporangia will
function as gametangia and one will empty its cellular contents into the
other.
---------- The final example in the
Lagenidiales is Lagenidium,
which is an aquatic fungus that may occur in wet soil. Many species in the genus are parasite and
some are saprophytes. One species
parasitizes the eggs of blue crab and lives on algae. It possesses a branched mycelium and is
holocarpic. During the asexual stage
of life cycle a zoospore lands on an alga and sends out a branched mycelium.
Septation occurs, which is wider
spread than in Myzocytium, and there is no rounding away of the
individual cells.
A sporangium develops on the
outside of the host from the cells of the mycelium.
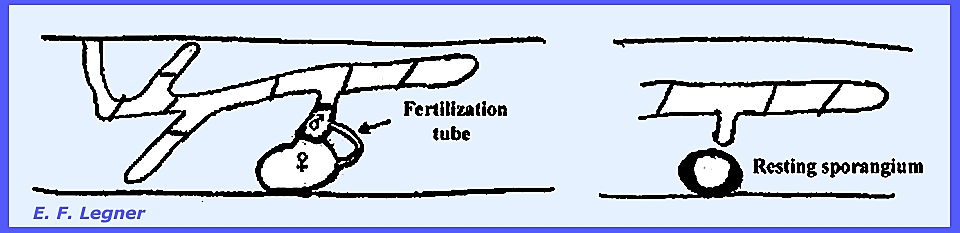
During the sexual phase one of the
septate sections of the myceliar cells rounds off into a female gametangium
and a male cell near to it sends out a fertilization tube. After transfer of protoplasm to the female
a resting sporangium is formed.
---------------------------------------- Please see
following plates for life cycles in the Lagenidiales: Zygomycota: Biflagellatae: Lagenidiales Plate
30 = Life Cycle -- Lagenidiales:
Olpidiopsis vexans Plate 31 = Life Cycle – Lagenidiales: Myzocytium
proliferum Plate 32 = Example Structures -- Lagenidiales: Lagenidium callinectes Plate 33 = Life Cycle – Lagenidiales: Lagenidium
spp. Plate 34 = Life Cycle – Saprolegniales: Saprolegnia
sp-1 Plate 81 = Life Cycle -- Lagenidium rabenhorstii. --------------------------------- The order Saprolegniales
or water molds, are
more typical of the Biflagellatae. Hyphae
are coarse (50 microns diam.) and very conspicuous. They attack a great variety of animal matter in the water, but
they are mainly saprophytes. Some,
such as Saprolegnia parasitica and Achlya flagellata, are parasites that attack fish and fish eggs. Other species are root and algae
parasites, e.g., Aphanomyces enteiches. Different authorities have variously drawn
the limits of this order, which varies from only one to three families,
etc. Only one family, the Saprolegniaceae,
will be presented here. The Genus Saprolegnia
is a typical representative of this order.
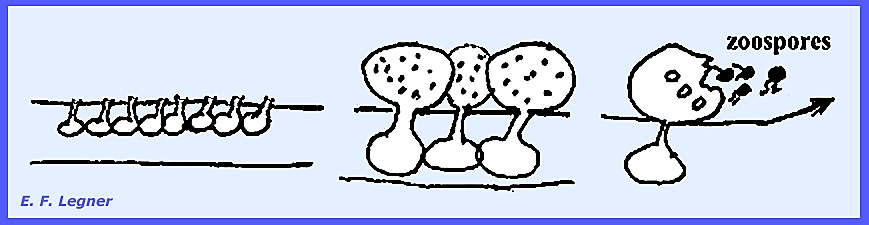
The species occur in great abundance in fresh water with most being
primarily saprophytic and a few parasitic on fish. The life cycle includes a branching mycelium that emanates from
the host (hemp seed) and forms zoosporangia on the tips of hyphae.
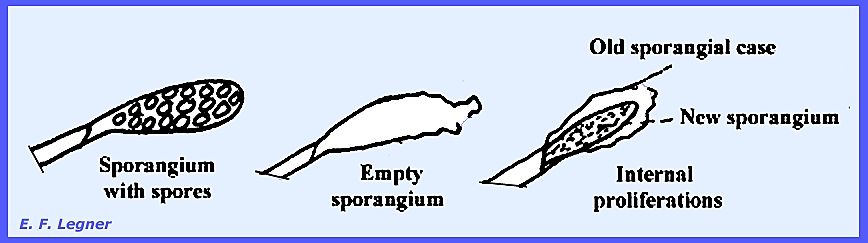
In the
asexual phase progressive cleavage delimits zoospores in the sporangium,
which is followed by internal proliferations.
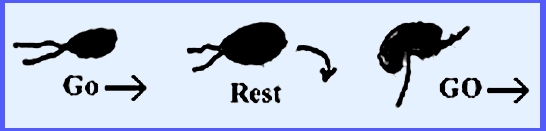
The zoospores are pear-shaped;
they swim in water and form a wall when on the host. The zoospores resume swimming but this
time their shape has become reniform.
In Diplanaticism there are two types of zoospores produced,
which have two swarming periods.
Occasionally
a hypha will swell up and the irregular-shaped tips (gemmae) break off and
give rise to a new mycelium. The
organism possesses a minor reproductive system.
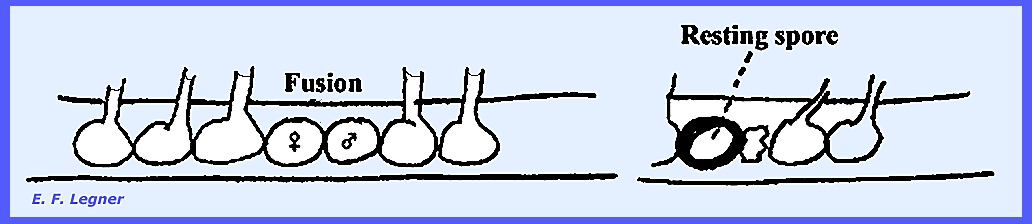
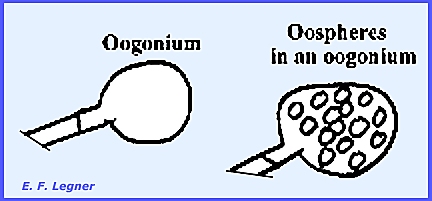
In the sexual
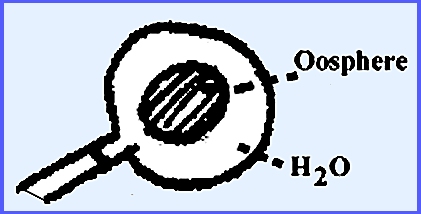
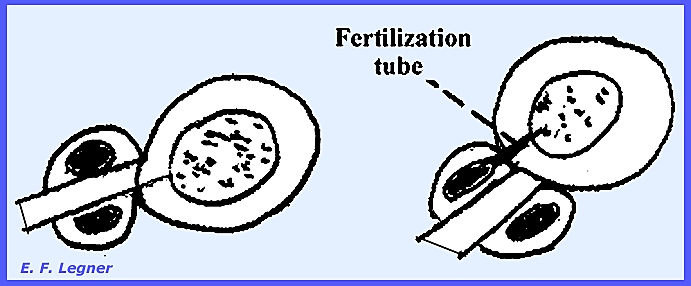
phase gametangia form on the ends of the hyphae and are called Oogonia. Oospheres (eggs) are delimited inside.
Antheridia
may form on another hypha or the same one.
These send fertilization tubes through the wall of the oogonium and
directly to each oosphere within.
Each oosphere develops into an oőspore after
fertilization.
The oöspores can germinate and the
tip may develop into a sporangium. If
oospores develop without fertilization they are called aboöspores (developed
parthenogenically). Review of the Saprolegniales The Asexual stage has a well-developed
mycelium, which consists of coarse, sparsely branching hyphae. Gemmae, zoosporangia are present and
pear-shaped zoospores are biflagellated at the apex and primary while the reniform zoospores are
secondary. In the Sexual stage there
is an oogonium with oospheres (eggs).
There are an antheridium, a fertilization tube and oosphores that give
rise to a hypha, which produces sporangia and send out zoospores. Variation in the Order Saprolegniales There are three types: Diplanetic, Monoplanetic and Aplanetic (=
two, one or no motile zoospores).
Example of the different types follows with some genera having both
types: Diplanetic: In Achlya there is no inner
proliferation of sporangia, and the sporangium arises from new growth below
the old one (later proliferation).
In Isoachlya there is both
a lateral and internal proliferation.
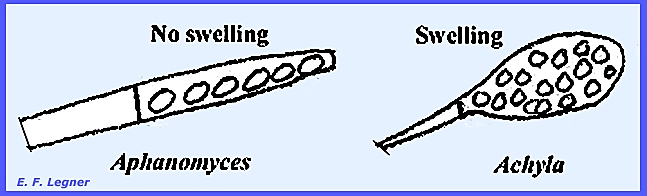
In Aphanomyces, Aplanes, Thraustotheca and Dictyuchus
there is no proliferation. Saprolegnia
and Isoachlya are also diplanetic.
In Achlya and Aphanomyces there is encystment in groups
at the mouth of the sporangium, and the reniform type follows and is the one
that swims. In Aphanomyces,
which is pathogenic on peas, the sporangium differs from Achlya.
Monoplanetic: In Thraustotheca the sporangia are
bulbous, the sporangia never open and the primary zoospores encyst right in
the sporangium. A breakdown of the
zoosporangial wall releases encysted primary zoospores and these give rise to
the reniform type.
In Dictyuchus after the
sporangia are formed, the zoospores become tightly packed inside in an
angular arrangement. Reniform spores
emerge from a hole in the side. A
deciduous sporangium is present.
Aplanetic: Aplanes has no swimming zoospores
and in Geolegnia there is an encystment of primary zoospores in the
sporangia. A germ tube is sent out
directly through the wall of the sporangium from each zoospore.
------------------------------------------- Sexual Phase Differences in the Saprolegniales In the Genera Aphanomyces, Dictyuchus
and Geolegnia there is a single oosphere in one oogonium and the whole
protoplast of the oogonium is used up in the production of the oosphere. Water lies between the oogonial wall and
the oosphere.
The majority of species are
monoecious where both reproductive structures are borne on the same
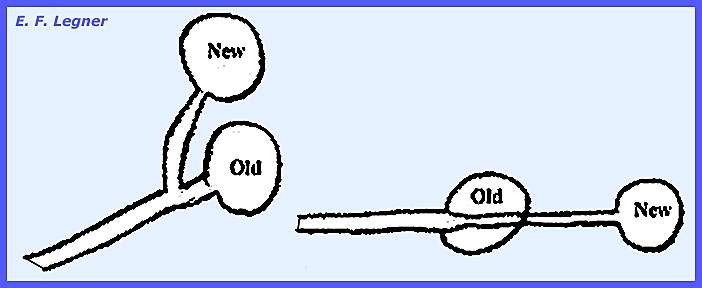
mycelium. However, some species, as
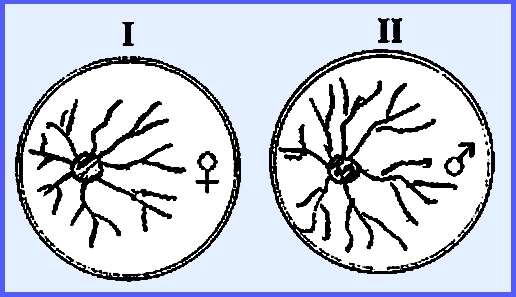
in Achlya, are dioecious. In the
diagram below when cultures I and II are separate no sexual structures are
formed. But when fluid of I (female)
is placed into II (male) it will cause the male thallus to produce antheridia
(Hormone A involved).
The male
produces another Hormone B, which causes the female to produce oogonia. Once the female has oogonia it produces
Hormone C that causes the male to develop an attraction. The male then produces Hormone D, which
causes delimitation of oospheres. ------------------------------------------- Please see the
following plates for life cycles in the Saprolegniales: Zygomycota: Biflagellatae: Saprolegniales Plate
35 = Example Structures – Saprolegniales: Saprolegnia sp2.
Plate 36
= Genera of Saprolegniaceae: Saprolegnia, Isoachlya, Achlya,
Aphanomyces, Aplanes, Thaustotheca,
Dictyuchus Plate
82 = Mature sporangium of Saprolegnia sp. Plate
83 = Dictyuchus sp.: Sporangium & zoospores. Plate
84 = Gemmae of Saprolegnia spp. Plate 85 = Saprolegnia litoralis: Terminal oogonium & Intercalary
oogonium. Plate
86 = Life Cycle -- Saprolegnia sp. -------------------- All species in the order Leptomitales
are saprophytic and aquatic. They
flourish in highly polluted waters.
Most are relatively unimportant save for their breaking-down
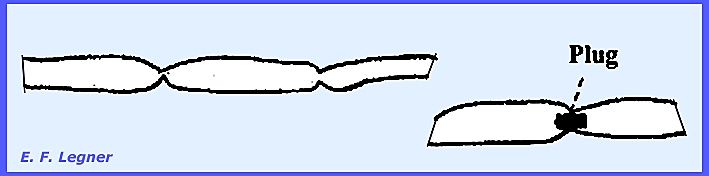
action. They possess a well developed
mycelium, which is coenocytic but always constricted. Plugs of celluin may be in the
constrictions.
Some species, cush as Rhipidium
and Sapromyces, may have an arbusculate mycelium. The following discussion on representative
genera will reveal the characteristics of this order. The Genus Leptomitus has constrictions
that occur at the branch points of the mycelium.
The genus cannot use sugar as a
carbon source, but instead uses organic acids and thus they are adapted to
polluted waters. There is no sexual stage
known. The tip of a mycelium branch
may turn into a sporangium, and septation may or may not occur before the
sporangium.
The species are also diplanetic (first pear-shaped followed by
reniform), and there is no proliferation although the next cell may function
as a sporangium.
The Genus Rhipidium has conspicuous
swellings in the sporangium and a tendency towards globular sporangia.
The zoospores are delimited in a
sporangium of the reniform type. No
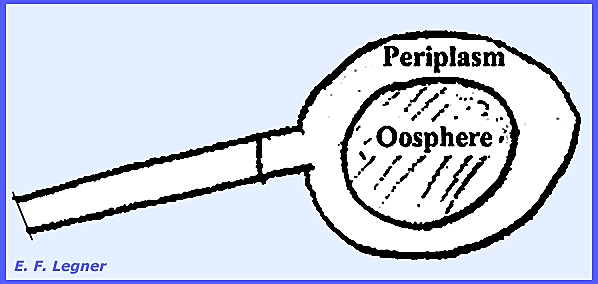
pear-shaped are ever formed in the sporangium (this in contrast to the Genus Thraustotheca). In the Genus Sapromyces there is a single
oosphere produced, and the protoplast is differentiated into two areas: an oosphere and a periplasm. The periplasm is left as an area between
the oogonium wall and the oosphere.
An antheridium is produced on the same thallus and gametangial
copulation takes place. There is an
arbusculate mycelium.
---------------------------------------- Please see the following for examples of Leptomitales: Zygomycota: Biflagellatae: Leptomitales Plate
87 = Leptomitales: Apodachlya
pyrifera & Rhipidium americanum. ---------------------------------------- The order Peronosporales
is the most advanced
order in the Biflagellatae. They are mostly terrestrial species, although
some are aquatic or semi-aquatic.
They are primarily parasites on higher plants, a considerable portion of
which are obligate parasites, such as Phytophthora that causes Downy mildews
and Pythium causing
White rusts. The probable evolutionary sequence of
families from lower to higher is Pythiaceae (damping-off fungi), Albuginaceae
(white rusts) and Peronosporaceae (downy mildews). Key characteristics of the group
is a well-developed coenocytic mycelium, and all produce globular oogonia and
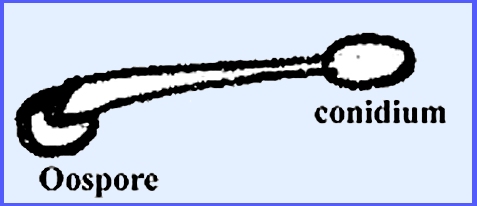
a single oosphere and periplasm.
Fertilization results in an oospore that is normally sculptured, the
latter formed from the collapsed portion of periplasm. The antheridia emit a fertilization tube
that persists. Sporangia are short
and wide and detachable in higher forms.
Detachable zoosporangia produce either conidia or sporangia, while the
hyphae that bear these produce either conidiophores or sporangiophores.
Zoospores are cut out usually after
detachment and they are released shortly afterwards (= indirect
germination). Under
different climatic conditions the zoospores may not be cut out in the conidia
and the latter simply germinate via a germ tube (= direct germination). These behaviors are characteristic of
highly evolved fungi and is shown in the Peronosporaceae. The order is mainly distinctions
of this order are the coenocytic mycelium that occurs mostly inside host
tissue. In the vegetative state
sporangia are present but conidia are in the more advanced forms. The zoospores that are produced from
sporangia are all reniform and in the more advanced forms the sporangia
germinate directly to form a new mycelium.
Sexual reproduction is remarkably uniform in the group. The oogonium always contains one oosphere
that is accompanied by periplasm. The
antheridium produces a fertilization tube that gives rise to an oospore,
which in turn gives rise to zoospores in similar ways. Key Families of
Peronosporales The Pythiaceae
are the most primitive
family in the order. Some lowly members thrive in aquatic habitats and they
often appear on hemp seed bait put out to trap organisms belonging to the
Saprolegniales. The higher forms tend
to be terrestrial. They are never obligatory
parasitic but rather facultatively saprophytic or parasitic. The Genus Pythium, causing “Damping-off”
and many other plant diseases, is abundant in soil and in some waters. They are low grade parasites that kill
tissue rapidly and then go on to live on the dead tissue either intra- or
intercellularly. They destroy the
hypocotyl region of seedlings or some may cause root rots. Because the sexual stage is so uniform,
vegetative features are the primary basis of separation of families Zoosporangia are produced from unspecialized hyphae and
zoospores develop from the vesicle that emanates from the sporangium.
Proliferations when they occur are
often lateral, but they may be internal as well by going through the old
sporangium.
The sexual
stages are typical for the order.
There is an oogonium with periplasm and an antheridium on the same or
different hyphae. After fertilization
the oospore is formed. The outer
layer of the spore is of decomposed periplasm. The oospore may bear sporangiospores directly or a hypha may
bear a sporangium on its end, which in turn produces zoospores. Phytophthora infestans causes “Late
Blight of Potato and
Tomato.” It is a facultative
saprophyte. In 1840 this species
caused disease in epidemic proportions in Europe, but was especially serious
in Ireland. It probably did not show
up earlier because of the mode of shipping where high temperatures over a
prolonged period in the holds of ships sterilized the potato tubers. By 1880 Bordeaux Mixture afforded partial
control. During the life cycle [see Plate 38] the fungus may
invade the potato tuber in addition to aerial portions. Mycelium grows on rotted tissues as a
saprophyte. The fungus is carried
over in the tuber and sporulation is on the young shoots. In the vegetative cycle mycelium
produces conidiophores, which emerge through stomata and are coarse with
diagnostic swellings at points where the conidia were attached. The conidia may do one of two things: either they may form zoospores through
indirect germination, or they may germinate directly without production of
zoospores (direct germination). The
direct penetration is through the cuticle whereas the indirect penetration is
through the stomata.
In the sexual cycle mating is
controlled by a compatibility factor: a heterothallic form or possibly a
facultative homothallic form. It is
not known if there are male strains and female strains in the oogonia and antheridia. The antheridium is borne at the base of
the oogonium and is “collar-like.” It
is believed that the oogonium punctures the antheridium and grows right
through it.
The life cycle of another species,
Pythium debaryanum, is shown in Plate 37. The Albuginaceae,
or “White Rusts” are generally not
as important as the other groups in the Peronosporales. Only a single genus, Albugo,
occurs. Albugo candida shows up as
white spots or blotches on plant leaves, which are caused by conidial
pustules [see Plate 39]. Plants are killed very slowly and
infection of inflorescences are serious.
Sepals and petals turn green and become hypertrophied. During asexual reproduction the mycelium
grows in intercellular spaces where it is coenocytic and sends out haustoria. Conidiophores are borne very close
together just under the epidermis.
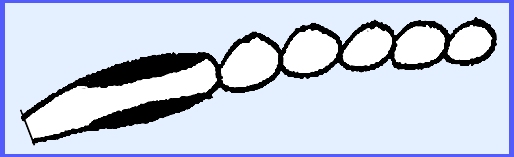
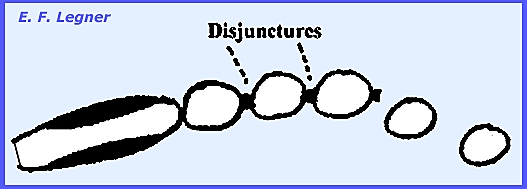
The conidia are catenulate.
Pressure from underneath breaks
the epidermis and spores form disjunctures followed by being released through
wind and rain primarily.
The conidia, functioning as
sporangia, form zoospores that penetrate indirectly. Sometimes in cases of low moisture and
high temperature germination may be direct where a germ tube is produced from
the conidium. Albugo is,
therefore, most prevalent in spring and autumn. During sexual reproduction the
mycelium, which produces conidiophores, forms multinucleate oogonia in
intercellular spaces. A single
oosphere is formed with only one nucleus.
Other nuclei disintegrate in the periplasm. The nucleus from the antheridium fuses with that of the
oosphere. The oospore that is formed
differentiates into a 3-layered wall.
Many oil droplets occur in the
center, which fuse to form one large oil droplet that is in a half-moon
shape. The diploid nucleus divides to
form many nuclei. The fertilization
tube becomes conspicuous and holds the oospore in position inside the empty
oogonium. A rest period will occur. Later the oospore wall cracks open and a
balloon-like vesicle emerges and delimits zoospores. The number of zoospores produced is more
than the number of nuclei before the rest period. Hence, it is believed that two nuclear divisions occur during the
resting phase. The Peronosporaceae,
or “Downy Mildews” are all obligate
parasites on angiosperms, causing great economic damage. They differ from the white rusts in that
the vegetative reproductive stage has distinctive branching conidiophores,
which grow out through the stomata (determinate). There are haustoria and intercellular mycelia with nuclei. Detachable sporangia occur that are
commonly called conidia. These may
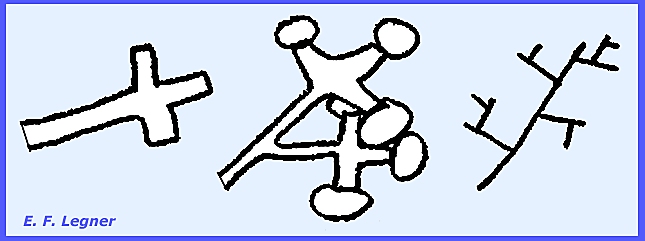
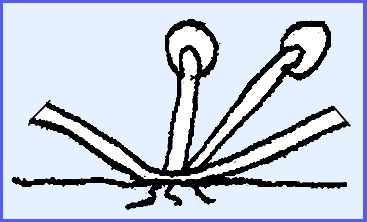
germinate directly or give rise to zoospores. The sporangia are determinate in growth. Plasmopara viticola causing “Downy Mildew of Grape.”
Is a fungus of wild grapes native to America [see Plate 40]. It is relatively unimportant on the native
American grapes, but in 1875 it was introduced into Europe where it proved to
be highly pathogenic on European grapes and practically ruined the wine
industry. In the 1880’s Bordeaux Mixture
(CuSO4 + CaO) was discovered accidentally as a remedy for
Downy Mildew. Vineyards that had been
treated with this mixture did not show a prevalence of Downy Mildew. The mixture proved useful against other
fungi as well. The pathogen, P. viticola,
forms mycelium in intercellular spaces with haustoria. It is rather coarse and irregular in shape
and the haustoria are larger than in Albugo.
Conidiophores grow out through the
stomata. They are tree-like and
monopodial (= single branch at a node).
The tips of the sterigmata are
rather blunt. The conidia readily
fall off and are carried away by rain.
They are determinate in growth.
Emergence is always through live host tissue and the area may commonly
be chlorotic. Conidia may function as
zoosporangia or germinate directly.
The sexual stage is similar to Albugo the only difference being that
the oospore germinates with a germ tube that bears a single conidium at its
apex.
-------------------------------------------- Considering other generaa in the Peronosporaceae
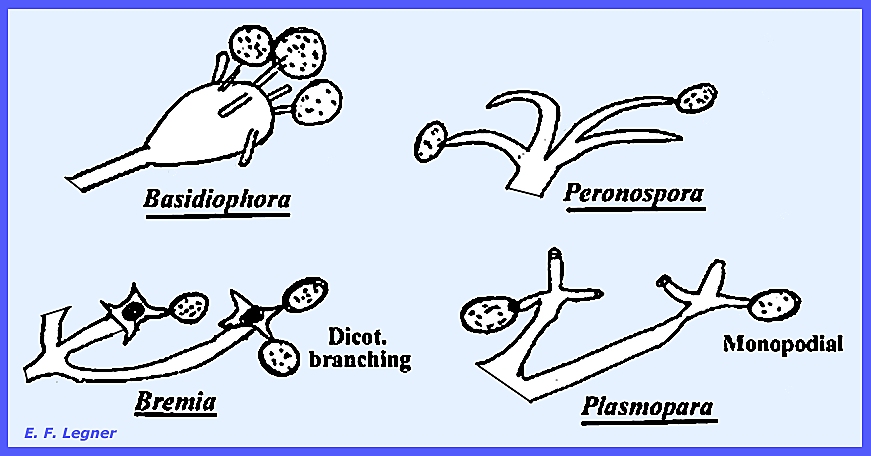
the largest and most complicated haustoria are found in the Genus Peronospora.
Almost all obligate parasites that
have mycelium also have haustoria.
This does not indicate that all haustoria-producing fungi are also
obligate parasites. Under favorable
conditions the conidia release zoospores in Plasmopara, but in Bremia
there is a greater tendency to germinate directly. Peronospora always germinates directly. The genera can also be distinguished by
that occurrence of conidia on distinctive conidiophores.
------------------------------------------- A review of the differences among
the three families of Peronosporales is as follows: Pythiaceae. Phytophthora possess haustoria
while Pythium does not. There
are no obligate parasites and conidiophores grow out through the
stomata. They have indeterminate
growth. Albuginaceae. Albugo are obligate parasites on
angiosperms. They have knob-like
haustoria without nuclei.
Conidiophores are club-shaped and are tightly packed in the
subepidermal layer. Conidia may
germinate directly or produce zoospores.
There is a single oosphere, periplasm and antheridium. Peronosporaceae are all obligate
parasites on angiosperms and differ from Albuginaceae in that the vegetative
reproductive stage has distinctive branching conidiophores that grow out
through the stomata. The haustoria
and intercellular mycelium do not have nuclei, and the detachable sporangia
may germinate directly or give rise to zoospores. The sporangiophores are
always determinate in growth. ----------------------------------------
Please see the following plates for life cycles and structures in the Peronosporales: Zygomycota: Biflagellatae:
Peronosporales Plate 37 = Life Cycle – Biflagellatae:
Peronosporales: Pythiaceae: Pythium
debaryanum Plate 38 = Life Cycle – Biflagellatae:
Peronosporales: Pythiaceae: Phytophthora
infestans Plate 39 = Life Cycle – Biflagellatae:
Peronosporales: Albuginaceae: Albugo candida Plate 40 = Life Cycle – Biflagellatae:
Peronosporales: Peronosporaceae: Plasmopara
viticola Plate 41 = Example Structures – Biflagellatae:
Peronosporales: Pythiaceae: Phytophthora
infestans Plate 42 = Example Structures – Biflagellatae:
Peronosporales: Albuginaceae: Albugo
spp. Plate 43 = Example Structures – Biflagellatae:
Peronosporales: Peronosporaceae: Plasmophora,
Bremia & Basidiophora. Plate 88 = Peronosporaceae haustoria: Peronospora ficariae, Plasmopara
pygmaea & Peronospora parasitica. Plate
89 = Life Cycle -- Pythium debaryanum. Plate
90 = Life Cycle -- Phytophthora infestans. Plate
91 = Sporangiophors in 5 genera of
Peronosporaceae. Plate
92 = Life Cycle -- Plasmopara viticola. Plate
93 = Life Cycle -- Albugo candida. Plate
94 = Oospores of 6 species of Albugo. ---------------------------------------- Aflagellatae
Three principal orders in the Aflagellatae
are Mucorales, Entomophthorales
and Zoopagales. The Mucorales are
for the most part saprophytic organisms.
Some species are extremely common and constitute a prominent element
in the “mold” population. Their
spores by often being abundant in the air readily contaminate any exposed
objects. These spores may germinate
and give rise to mycelia on suitable substrates. The order plays a significant role in the decomposition of
organic waste. However, they also
cause extensive damage through spoilage of food and they are a nuisance as
laboratory contaminants. A few
species act as low-grade parasites and may occasionally cause destructive
rots of living plant structures. Rhizopus
causes a soft rot of vegetables and fruits after harvest, especially sweet
potatoes, white potatoes, strawberries, plums, etc. Uncommonly lung and ear infections in humans by some species
have been reported. Some mucors are
parasites on other mucors and various members of the order may be found as
soil inhabitants, and certain ones called “coprophilous” are ordinarily
encountered growing on dung. Among
the latter the most spectacular is Pilobolus. Many species form a conspicuous
cottony mycelium, which is coenocytic when young but as age progresses often
develops numerous crosswalls. There
are very interesting specialization of vegetative hyphae in some genera,
e.g., Absidia and Rhizopus.
During vegetative reproduction sporangia may be formed on
sporangiophores that may be branched or unbranched. Non-motile spores (sporangiospores) are borne in such sporangial
sacs. Some species bear diminutive
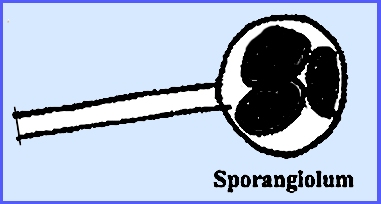
deciduous sporangia called sporangiola. The merosporangium
represents a peculiar sporangial type.
Some of the higher evolved species do not produce sporangia at all but
rather bear conidia that are believed to be derived from monosporous
sporangia. Chlamydospores are frequently
encountered and in some species these are very abundant. During sexual reproduction zygospores appear
following the union of gametangia that are frequently of almost equal size,
although in a few cases the size difference may be pronounced. Many species are heterothallic. The Genus Rhizopus, which includes the
black bread molds, serves as a typical example of the order. These fungi decompose soil and organic
matter and some are low-grade pathogens but may incite destructive rots of
fruits and vegetables. Some species
are animal pathogens causing eye and ear infections. The mycelium grows rapidly and is coenocytic,
coarse, cottony and profusely branching The hyphae have crosswalls in older
mycelia and septa almost always yield multinucleate cells. There are no septal pores, which is
different from higher true fungi. Stolons and a system of rhizoids
form at the point of contact with the substrate.
During vegetative reproduction there is growth at the junctures of
rhizoids, which are sporangiophores, that later bear sporangia.
A membrane is formed by the fusion
of vacuoles and later a fully formed wall is laid down. The basal portion of the sporangium is now
the columella, and progressive cleavage delimits aplanospores, which are wind
disseminated.
During sexual reproduction a very
thick-walled structure the zygospore is formed by the fusion of two
gametangia. ------------------------- The life
cycle of Rhizopus nigricans further
serves to characterize the genus [see Plate 44]. Zygospore formation is rarely seen in this
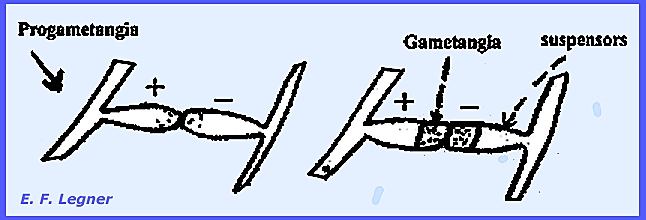
species because of heterothallism, which involves Plus and Minus strains. Progametangia
meet and change into gametangia and suspensors.
Fusion occurs
to produce a thick-walled, dark-colored zygospore.
Commonly one of the suspensors
swells up markedly.
Occasionally the gametangia will
develop thick walls without fusion.
These are azygospores
that function similarly as zygospores.
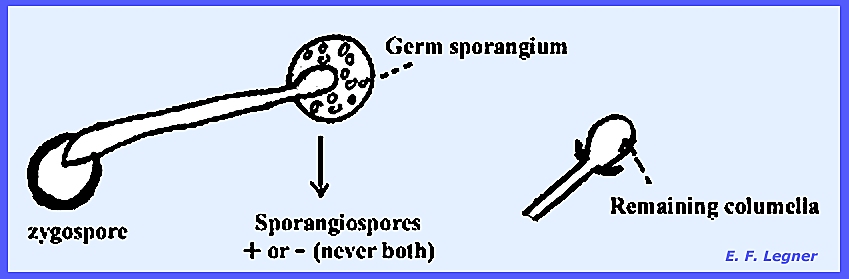
The zygospores eventually
germinate but they are rarely seen to do so.
A hypha grows out of the zygospore that terminates in a columellate or
“germ
sporangium.” The columella
will remain after the sporangium is shed.
--------------------------------------- Families and Genera of Mucorales [Also
see <Mucorales Key>] Five families are discussed as
representative of the Mucorales: Mucoraceae, Pilobolaceae, Piptocephalodaceae, Thaminidiaceae,
Cunninghamellaceae. Mucoraceae. --
In the Genus Rhizopus stolons and rhizoids occur at the point where
the stolon touches the substrate.
Sporangiophores arise at the junctions with rhizoids.
Some species like Rhizopus
nigricans are heterothallic while other species may show
homothallism. There is a well-developed
columella at the apex of a sporangiophore, and the wall of the sporangium is
quite delicate. Absidia is one of the
principal genera of the Mucorales encountered in the soil. The mycelium is more delicate than Rhizopus
and stolons and rhizoids have patterns that differ from Rhizopus. Rhizoids are inconspicuous and attached at
the apex of an extension.
Sporangiophores arise on the arch, singly or in small groups. The pear-shaped sporangia are quite
delicate.
During the sexual stage of Absidia
appendages are formed on the suspensors (either one or both suspensors bear
appendages). They are called “coiled appendages”
and emanate from one or both suspensors to form a basket-like structure. In Circinella’s vegetative
stage sporangiophores are long with short side branches that curl. Curling is called “circinate”
Sporangia are columellate and are
borne on the “curl.” Some species
have a mate to the circinate structure.
The sexual stage of Circinella is similar to Rhizopus.
Mucor is the largest genus
in the Mucorales. The species have a rapidly
growing mycelium and all are saprophytes and very common in the soil. There are homo- and heterothallic
species. The vegetative stage is
similar to Rhizopus but the sporangiophores are taller. There are no stolons or rhizoids (Absidia
and Rhizopus are the only two genera producing these structures in the
Mucorales). Sporangiophores may arise
anywhere and may be branched or unbranched. Syzygites (Sporodinia) are parasites of fleshy fungi (gill and
pore fungi). They are rapidly growing,
have a slightly yellowish mycelium, which forms sporangiophores in
abundance. The sporangiophores are
dichotomously branched.
The sporangia have columellae and each
sporangium contains a few sporangiospores.
The sporangiospores stick on the outside of the columella after
disintegration of the sporangial wall.
These species are homothallic.
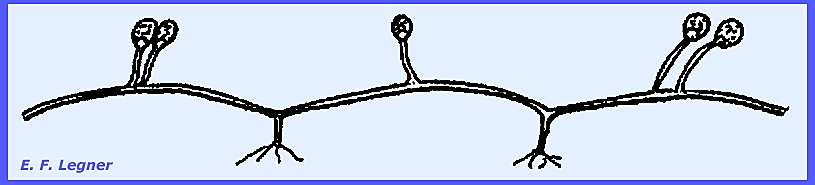
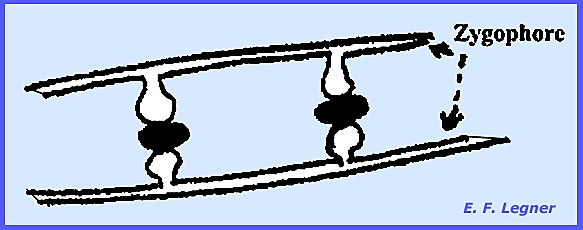
Gametangia develop on specialized hyphae called zygophores (=
stiffened hyphae).
In Phycomyces the sexual stage
is distinctive but very rare.
Suspensors form outgrowths, as in the case of Absidia. These are spine-like structures.
There are particularly large sporangiophores
coming from undifferentiated hyphae.
They are strong and coarse and yellowish-green when young. The sporangium is columellate and the
whole structure is sensitively phototropic.
Up to this point all of the
Mucorales have had a well-developed columella. ------------------------ Pilobolaceae.
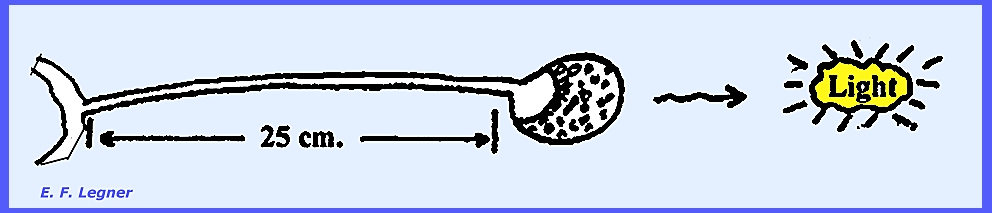
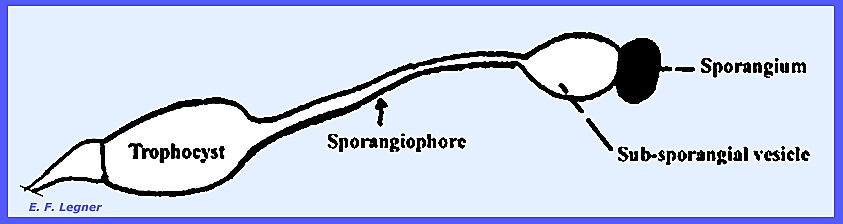
-- Pilobolus is a coprophilous genus where
the mycelium grows rapidly on dung and produces characteristic
sporangiophores. There is a bulbous
cell at the base of a sporangiophore called a Trophocyst or “Basal Bulb”. A subsporangial vesicle occurs at
the base of the sporangium, and a columella exists in the sporangium. A cap is produced, which is coal black.
There is a forcible
discharge of the sporangium where the sporangia are shot off. The sporangium lands with cap up; it
flattens out into a cushion and adheres to the substrate by a sticky
substance. In a phototropic response
the sporangium points eastward toward the rising sun. This orientation is quick and
remarkable. Spores land on the grass
after being discharged from the dung.
Herbaceous animals may ingest the sporangia and the spores are
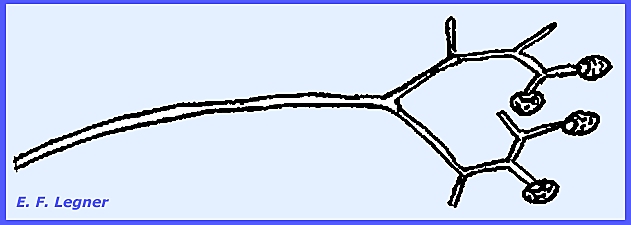
modified in the digestive tract to later germinate in the dung. --------------------------- Piptocephalodaceae.
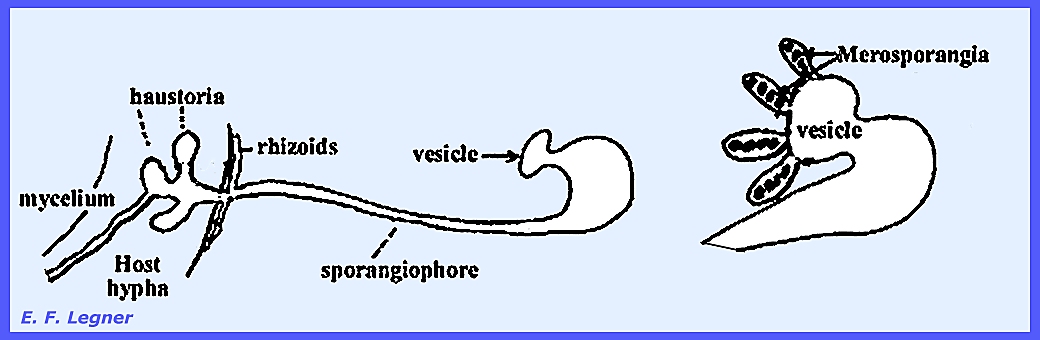
-- Syncephalis
parasitizes other Mucorales, which is rare for parasites to be found
in the same order as the host. The
mycelium grows closely attached to the host and sends out haustoria. They are high-type parasites and probably
obligate. The sporangiophores are
highly distinctive as is also the case with the sporangia. Rhizoids and haustoria anchor the
sporangiophore to the host hypha. A
vesicle appears at the apex of the sporangiophore and finger-like structures
grow out of the vesicle.
There is no columella and spores
are arranged in a single row. The
sporangia here are called merosporangia. In the Genus Piptocephalis dichotomous
branching of the sporangiophore occurs giving a broom-like effect. -------------------- Thamnidiaceae.
-- The Genus Thamnidium
has unbranched sporangiophores with columellate sporangia.
A different kind of sporangium
originates from another profusely branched sporangiophore, which is called a sporangiola. These do not possess a columella and are
deciduous. There are few
sporangiospores in the sporangiola.
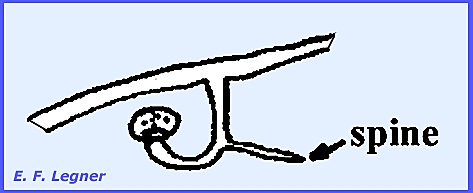
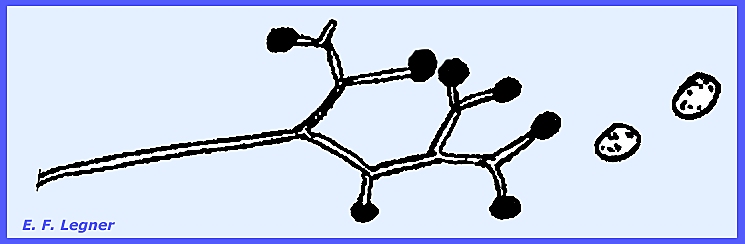
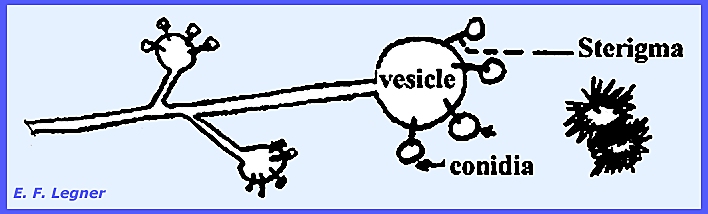
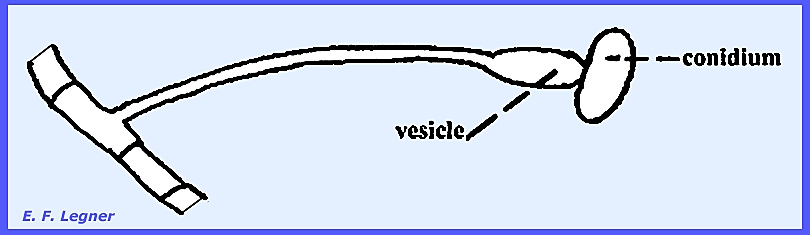
-------------------------- Cunninghamellaceae.
—Cunninghamella is
a common saprophytic form in the soil and decaying debris. There is nothing peculiar about the sexual
stage. A vesicle is produced on
sporangium-like structures and sterigmata emanate from the vesicle and
conidia are formed on them. Terminal
portions release conidia first. In
some species the conidia bear spines.
Essentially each conidium is a one-spored sporangiolum. The wall of the spore is contiguous with
that of the conidium or sporangium.
---------------------------- Please see the following plates for
life cycles and structures in the Mucorales: Zygomycota: Aflagellatae: Mucorales Plate 44 = Life Cycle – Aflagellatae:
Mucorales: Rhizopus nigricans Plate
45 = Example Structures – Aflagellatae: Mucorales: Absidia, Circinella, Cunninghamella, Mucor, Philobolus, Phycomyces, Rhizopus, Syncephalis, Syzygites, Thamnidium Plate 46 = Aflagellateae: Mucorales: Distinction of Genera in The
Mucorales: Absidia, Circinella, Cunninghamella, Mucor,
Philobolus, Phycomyces, Rhizopus, Syncephalis, Sygygites, Thamnidium Plate
95 = Stages of evolution of sporangium to a
conidium. Plate
96 = Life Cycle -- Rhizopus nigricans. Plate
97 = Zygospores of Mucorales. Plate
98 = Zygophore & Sporangiophore
formation: Sporodinia grandis. Plate
99 = Sporangial apparatus: Philobolus longipes. -------------------------------- Entomophthorales are represented here by six genera: Entomophthora, Massospora, Completoria,
Ancylistes, Conidiobolus and Basidiobolus. The distinction among three of these
genera may be viewed in Plate 49 This order is more advanced than the
Mucorales. Aflagellate forms produce
conidia, which in most cases are forceably discharged (except in the Genus Massospora). Spores are not formed until the conidia
are discharged. The majority of
species are parasitic on insects but they are not obligate parasites and
might best be considered facultative saprophytes. The mycelium is irregularly septate (cells irregular in
length), and septa occur in young hyphae.
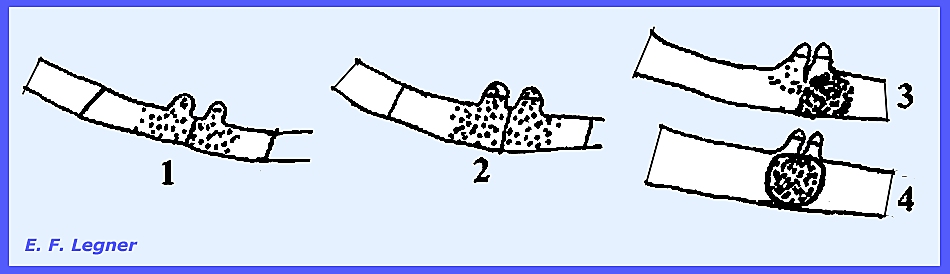
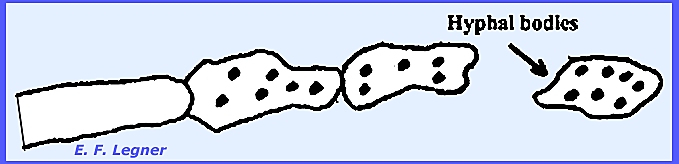
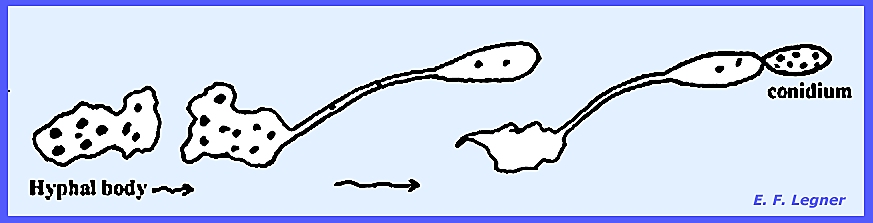
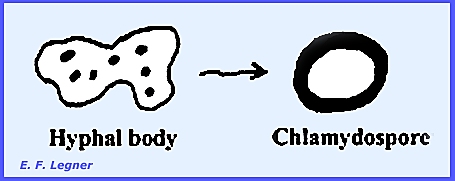
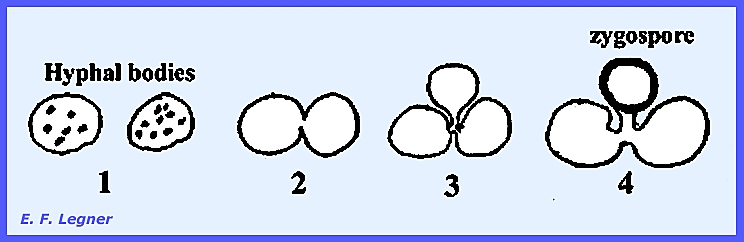
Hyphal
bodies form when cells of the mycelium disarticulate and pull
apart. Sexual resting spores, or
zygospores occur, and vegetative resting spores are called chlamydospores.
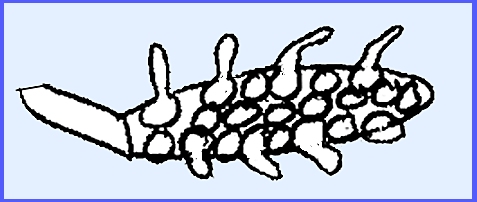
The Genus Basidiobolus is coprophilous on the dung of amphibians
and reptiles. The mycelium is rapid in
development, and the septate hyphae may form hyphal bodies. Conidiophores arise from cells of the
mycelium and there is a forcible discharge of the conidia.
Upon discharge the conidia land on vegetation where beetles may ingest
them. Later frogs and snakes may
consume the beetles. The conidia give
rise to internally formed spores tht function as sporangia. The wall of a sporangium breaks down
releasing sporangiospores in the dung.
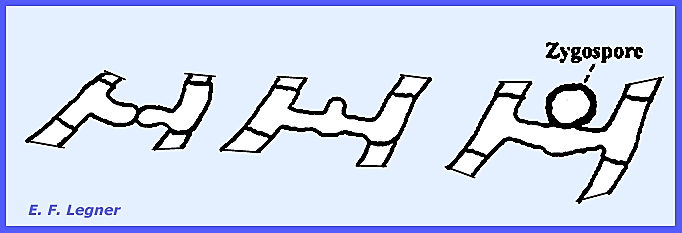
During the sexual stage, large thick-walled zygospores are formed in
abundance at intervals along the hyphae.
The genus is readily distinguished by the presence of two msall
projections (-= "beaks"
or "rabbit ears.")
These "beaks" form when two processes extend from adjacent
cells in the mycelium. A crosswall forms
across each process and fusion results in a manner not unlike earlier
organisms studied. The fusion takes
place inside the gametangium which in this case is the hyphal cell.
-------------------------
In the Genus Entomophthora all species are parasites on insects, and
infection is always fatal. As
epidemics occur frequently the genus is thought to be important in the natural
control of insects. This is the most
important genus in the order as it attacks many kinds of insects. Once infection is accomplished, an insect
rarely survives attack by one of these fungi..
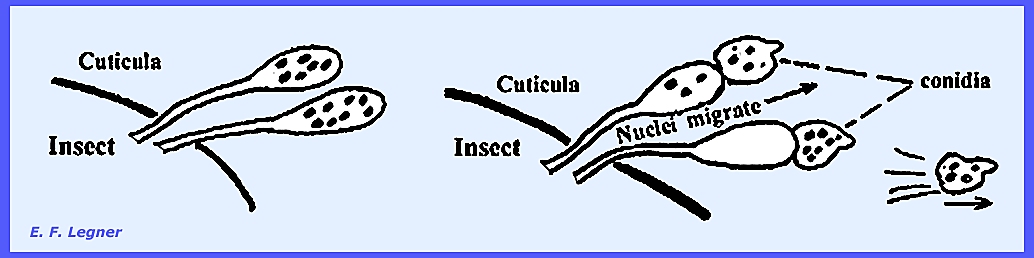
The mycelium of Entomophthora develops throughout the insect
body. The hyphae are coarse,
irregularly septate and may form irregularly shaped hyphal bodies.
Conidiophores grow out to the outside of the insect after death to
form a "halo" around the dead fly on the window pane or other
surface.
The conidium has a small beak and is forcibly discharged. It then sticks to surrounding objects,
which may include another fly. If the
conidium fails to land on an insect it will produce another conidium that is
again forcibly discharged, and this may be repeated several times until vigor
is lost.
Every
conidiophore takes its origin from a hyphal body in some species. It may produce characteristic
conidia. Germination of conidia is
direct.
Chlamydospores
are present in some species and form when a hyphal body may simply round
up to form a thick wall and become a resting spore.
During
Zygospore
formation the entire hyphal cell acts as a gametangium.
Hyphal
bodies may also behave as gametangia.
-----------------------------
The Genus Massospora also contains entomophagous forms, one
species attacking the 17-year Locust.
It differs from Entomophthora in that it does not forcibly
dicharge conidia [see Plate 49]. Conidia are
found inside the insect's posterior portions. Posterior segments drop off the insect and the conidia are
diseminated during the final movements of the insect before death. -------------------------------- Please see
the following plates for life cycles and structures in the Entomophthorales: Zygomycota: Aflagellatae: Entomophthorales Plate 47 = Life Cycle – Aflagellatae: Entomophthorales: Basidiobolus sp. Plate 48 = Life Cycle – Aflagellatae:
Entomophthorales: Entomophthora sp. Plate 49 = Example Structures: Aflagellatae,
Entomophthorales: Basidiobolus, Entomophthora, Massospora Plate
100 = Structures of Entomophthora muscae. Plate
101 = Structures of Entomophthora
sepulchralis. -------------------------------- Zoopagales is one of the most highly specialized and
advanced groups in the Zygomycota. They
parasitize amoebae, nematodes and insect larvae in the soil. The mycelium is fine and septate and
conidia are produced in all of the species.
Zygospores are present also.
They may serve to regulate the populations of the small forms of
animal life in the soil. = = = = = = = = = = = = = = |