File:
<bushfly.htm> GENERAL INDEX [Navigate to MAIN MENU
]
|
CONTROL OF
THE BUSH FLY & Other Diptera in The Kwajalein
Atoll, Marshall Islands(1) E. F. Legner (2), B. B. Sugerman (3), Hyo-sok Yu (4), and H. Lum (5) (Contacts) Please CLICK on underlined figures, tables & footnotes to view details] GO TO ALL: Bio-Control
Cases SUMMARY An investigation into the
biological-integrated control of pestiferous flies in Kwajalein Atoll,
Marshall Islands revealed 6 principal species in the families Calliphoridae,
Sarcophagidae and Muscidae, among which some were successfully reduced to
non-annoyance population levels in 18 months. Musca vetustissima Walker,
of the Musca sorbens complex, was satisfactorily reduced on some
islands through habitat reduction, the introduction of natural enemies and
baiting after 21 months. The history, biology and control of M.
vetustissima is reviewed and future efforts outlined for the importation
of natural enemies. Possibilities for resistance to insecticides are
indicated and precautions recommended for the effective long term use of
poisoned baits. The potential invasion of M. sorbens in North America
is discussed. (7) INTRODUCTION Pestiferous flies in the Marshall Islands provide a
classic example of the adaptation of invading noxious insects to a salubrious
tropical climatic area. With nearly perfect temperature-humidity conditions
for their development, an abundance of carbohydrate and protein-rich wastes
in the form of garbage and excreta provided by man and his animals, and a
general absence of effective natural enemies, several species were able to
reach maximum numbers. There are principally 4 types
of pestiferous flies in Kwajalein atoll of the Marshall Islands, with the
Australian bush fly, Musca vetustissima Walker of the Musca sorbens
complex, being by far the most pestiferous species (FIG. I). The common house fly, Musca demestica L., of
lesser importance, frequents houses and is attracted to food in recreation
areas. The remaining 2 types are the Calliphoridae [Chrysomya megacephala
(Fab.), and Phaenicia cuprina (Wiedemann)) , and the Sarcophagidae [ Parasarcophaga
misera (Walker), and Phytosarcophaga gressitti Hall and Bohart).
These latter species are abundant around garbage disposal sites and wherever
rotting meat and I decaying fish are available. The Calliphoridae are copper
to bluish-green, sometimes blue, on the thorax making them easily
distinguishable from the large striped gray Sarcophagidae. Both of these
flies differ from the common house fly and the bush fly in being more sluggish
and noisy and by their general avoidance of humans. Because residents do not
a distinguish different kinds of flies, these latter 2 types are often blamed
as nuisances when in fact they may be considered to fulfill a useful role in
the biodegradation of garbage and rotting a meat. PROCEDURE Studies were begun in April 1971 at the request of the
United States Army and the United States Trust Territory of the Pacific, to
investigate fly abatement possibilities in Kwajalein Atoll, Marshall Islands.
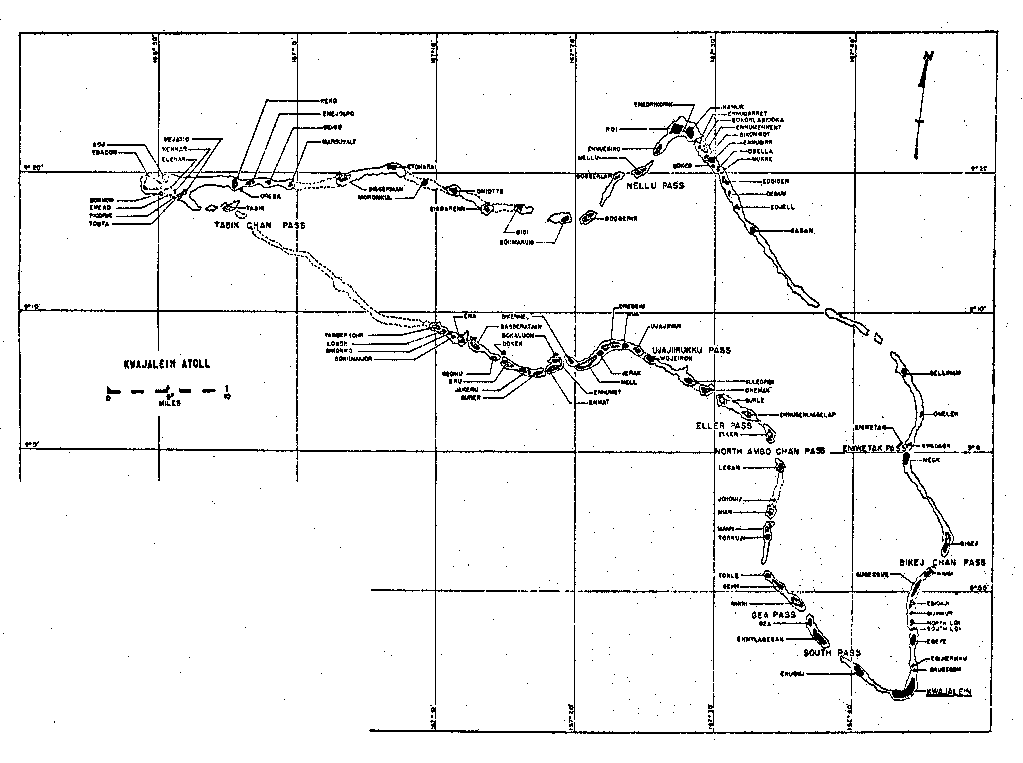
The atoll, the largest in the world, is about 120 Km long by an average 23 Km
wide and contains 93 islets grouped roughly between 166-167° E. longitude and
8-9° N. latitude in an irregular ring around a central lagoon (Bryan 1972 and
FIG.
2).
Kwajalein, the largest island, lies at the extreme south of the atoll
(FIG. 2 , 3 & 5). An initial assessment of the problem using standard
sampling methods to locate fly breeding sources and identify associated
natural enemy fauna, resulted in the expedient implementation of breeding
source reduction to reduce the house fly, Musca domestica L., and both the
Calliphoridae and Sarcophagidae to inconspicuous levels, which largely
involved slight modifications of refuse disposal sites to disfavor fly
breeding. By November 1972, these simple measures resulted in an estimated
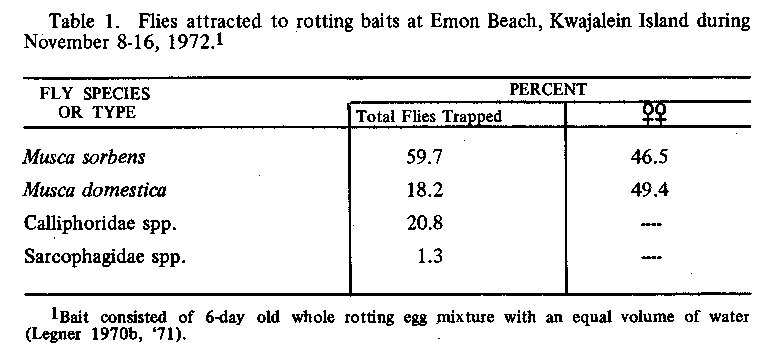
1/3rd reduction of total flies concentrating around beaches and other
recreational areas (see Table 1 for proportion of
fly species trapped). Because the house fly especially enters dwellings, its reduction was desirable for the
general health of the community, and fly annoyances indoors diminished. Thorough surveys of breeding sites and
natural enemy complexes revealed that Musca vetustissima reduction would not be quickly forthcoming,
however. A schedule of importation for natural enemies was begun in 1971 and
other integrated control approaches were investigated: eg. baiting and
breeding habitat manipulation. The
following account of this, the principal problem fly species in the atoll,
reviews bush fly history and biology, at the same time pointing out the
magnitude of the biological, technological and political difficulties
involved in its permanent reduction. RESULTS AND DISCUSSION Bush
Fly. Musca vetustissima Walker of the Musca
sorbens 1830 complex Origin and Habits This group of flies is known as the bazaar fly in North
Africa, a housefly in India, and the bush fly in Australia (Yu 1971). Musca
sorbens was first described from Sierra Leone in West Africa in 1830
(Wiedemann 1830) where it is a notorious nuisance to man and animals. The
flies are attracted to wounds, sores, and skin lesions, searching for any
possible food sources such as blood and other exudations. Although not a
biting species, its habits of being able to transmit eye diseases, entheric
infections, pathogenic bacteria and helminth eggs make it one of our most
important and dangerous public health insects (Bell 1969; Buston and Hopkins
1927; Gaud and Faure 1951; Gaud et al. 1954; Greenberg 1971; Hafez and Attia
1958; Lamborn 1937; McGuire and Durant 1957; Patton 1920, '22, '23, '26, '32,
'33; Thomson and Lamborn 1934). The bush fly has spread through a major portion of the Old
World, Africa and parts of Asia (Awati 1920; Gaud and Faure 1951; Hafez and
Attia 1958; Hughes 1968; Jack 1931; Lewis 1954; Meng and Winfield 1944;
Norris 1966; Patton 1920, '23, '26; Roy and Siddoils 1940; Van Emden 1965).
In Oceania its distribution is in AustraIia (Norris 1966; Paterson and Norris
1970); New Guinea (Paterson and Norris 1970); Samoa (Huxton and Hopkins
1927); Guam (Harris and Down 1946, Hohart and Gressitt 1951); and the
Marshall Islands (Hohart and Gressitt 1951 )0 In Hawaii it was first reported
by Joyce in 1950. Since then Hardy (1952) listed it in the Catalog of
Hawaiian Diptera, and Wilton (1963) reported its predilection for dog excrement. Annoyance by the bush fly increased in
the 1960's when it was incriminated as a potential vector of Beta-haemolytic
streptococci in an eipdemic of acute glomerulonephritis (Bell 1969). Taxonomy. -- Paterson and Norris (1970)
identified 3 biological forms in the M. sorbens complex. These are
distinguished by morphometric differences and reproductive isolation in the
laboratory, and consist of the African male broad-frons and narrow-frons
forms, and the Australian narrow-frons form.
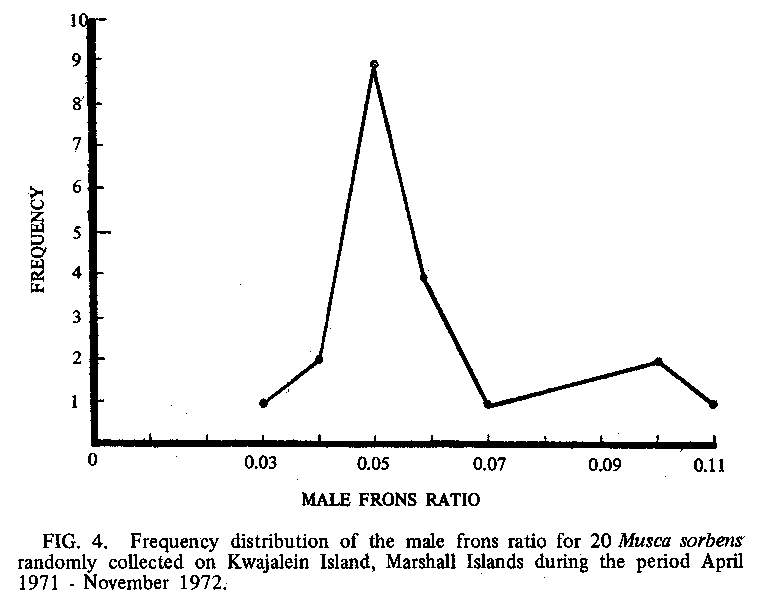
Examination of morphometric measurements of 220 randomly
selected adult males collected on Kwajalein Island during the period April
1971 -November 1972 showed an average frons ratio (6) of 0.0590 (range 0.0341 -0.1052; SD = 0.0182), which was
close to that of both the African and Australian narrow-frons forms. However,
the frequency distribution showed 2 distinctive peaks representing the
narrow- and broad-frons in the ratio of about 4: I (FIG 4). Also, a count of the parafrontal hairs of
20 random females (Paterson and Norris 1970) gave similar results. Thus, 2
forms may be sympatric in Kwajalein, which has been shown possible from other
areas (Paterson and Norris 1970). Breeding Habits. -- The principal breeding sites of the bush fly vary according to
different geographical localities.
However, human excrement is regarded by many authorities as a
preferred site when this is available (Bohar,t and Gressitt 1954; Gaud et al.
1954; Hafez and Attia 1958; Harris and Down 1946; Norris 1966; Patton 1933;
Peffly 1953a, b; Roy and Siddons 1940; Sabrosky 1952). The bush fly also breeds in fresh
droppings of a number of larger animals, such as cattle (Bohart and Gressitt
1951; Hafez and Attia 1958; Hughes and Walker 1970; Jack 1931; Meng and
Winfield 1944; Pat ton 1922, '36; Peffly 1953b); horses (Bohart and Gressitt
1951; Lewis 1954; Pat ton 1922, '36); dogs (Meng and Winfield 1944; Norris
1966; Pat ton 1922; Wilton 1963); and pigs (Bohart and Gressitt 1951; Meng
and Winfield 1938; Smirov 1940; Hafez and Attia 1958). Sheep feces (Norris
1966), and goat and cat feces (Yu 1971) are also suitable. Poultry excrement will not support this
fly's development (Yu 1971). Other
breeding sources of much lesser importance but which are capable of producing
small numbers of bush fly are carcasses of animals, decaying vegetable,
matter and garbage (Bevan 1926, Norris 1966, Pat ton 1922, Yu 1971). Recent studies by Yu (1971) in Hawaii show
that dog feces are the most important breeding medium on Oahu. He concluded that dung of cows, horses,
pigs and goats are of minor importance in rural areas where dog feces are not
readily available. Development. -- The average developmental period in Hawaii from eggs to adults is
10 days with an average of 76 flies emerging per field sample of dog feces.
On Kwajalein developmental time probably averages 8 days with over 100 flies
emerging from one sample of dog feces. High precipitation may prevent the fly
from full activity and breeding (Yu 1971).
In the Kwajalein atoll there is little reason to doubt
that a substantial portion of the main density of Musca vetustissima
emanates from dog and human feces.
However, extensive inspection of pig droppings in the bush of 10
widely separated islets revealed high numbers of larvae (over 100 per
dropping), making this dung, as in Guam (Bohart and Gressitt 1951 ), a
primary breeding source in the atoll. Pigs that are corralled on soil or
concrete slabs concentrate and trample their droppings making them less
suitable breeding sites. In such situations flies were only able to complete
their development along the periphery of corrals. Coconut husks placed under pigs in corrals results in the
production of greater numbers of flies by reducing the effectiveness of
trampling. These conclusions were reached after comparing quantitative
samples taken over a period of 2 years from the respective breeding habitats.
Garbage was not found to breed M. vetustissima,
although admittedly a very low percentage of the adult population could
originate there judging from reports elsewhere. Nevertheless, this medium is
certainly not responsible for producing more than 2% of the adult densities
observed in the atoll. Control Efforts and Evaluation Worldwide. -- Successful partial control of
bush fly has been achieved only in Hawaii through a combination of the
elimination of breeding sites, principally dog droppings, and the activities
of parasitic and predatory insects introduced earlier for biological control
of other fly species, e.g., Musca domestica (Legner 1978 ). The density of-bush fly
varies in different climatic zones in Hawaii, but the importance of this fly
is minimal compared to Kwajalein. At times hymenopterous parasites have been
found to parasitize over 95% of flies sampled in the Waikiki area (H. Yu,
unpublished data). Other parts of Oceania were either not suitable for the
maximum effectiveness of known parasitic species ( e.g. Australia) or the
principal breeding habitats were not attractive to the natural enemies.
Therefore, in Australia a concerted effort is being made to secure scavenger
and predatory insects from Africa that would be effective in the principal
unmanageable fly producing source, cattle and sheep dung (Bomemissza 1970; G.
F. Bomemissza, personal communication). For example, insectary studies on Onthophagus
gazella F. (Coleoptera: Scarabaeidae) showed 80-100% reduction of M.
vetustissima emergence
from cattle dung (Bomemissza 1970). Surviving fly larvae gave rise to small,
stunted adults of reduced reproductive capability. Kwajalein Atoll. -- Continuing integrated fly control begun in 1971 has now reached a
level of partial success. Initial surveys for natural enemies of M.
vetustissima in April-May 1971 revealed the presence of 4 scavenger and
predatory insects, the histerid Carcinops troglodytes Erichson, the
nitidulid Carpophilus pilosellus Motschulsky, the tenebrionid Alphitobius
diaperinus (penzer), and the dermapteran Labidura riparia
(pallas). Following our recommendations in May 1971, dogs were significantly
reduced and all privies were reconstructed or improved on one island,
Ebeye Dogs were reduced or tethered
on Kwajalein Island and refuse fish, etc., disposed of thoroughly on
l1leginni (an island that experienced outbreaks of Sarcophagidae in 1970 (FIG. 2), and other islands with American residents. Continuing importations of natural enemies
were made throughout the atoll with establishment of some species verified in
November 1972 (Table 2). The average density of M. vetustissima on Ebeye was,
subsequently, considerably reduced by November 1972 from an estimated 8.5
flies attracted to the face per minute!, to less than 0.5 flies per minute, a
reduction readily recognized by the inhabitants (7). The single most
important cause appeared to be the partial elimination of breeding sources,
with natural enemies playing a secondary role The M. vetustissima
density at Roi-Namur (estimated 2.5 flies attracted per minute), in the
norther apex of the atoll, and Ennylabegan (6.0 flies per minute), in the
southwest, did not substantially improve following the natural enemy
introductions and recommendations for reducing breeding habitats. However, this is thought to be due to an
increase in the number of human inhabitants, dogs and pigs on Ennylabegan and
on Ennubira to the southeast of Roi-Namur (Fig. 2). Flies apparently invade islands adjacent to their breeding
origin at a very slow rate as evidenced by a lack of recovery on Kwajalein of
dyed flies that were released on Ebeye and Ennylabegan in November, 1973.
However, Australian studies support the fact that M. vetustissima possesses rapid,
long-distance migrating behavior, which is aided by windstorms (Hughes 1970,
Norris 1966). Even though mark and recapture experiments were negative, there
is probably continuous slow invasion of M. vetustissima over a proposed route from Ennubira to
Roi-Namur; and from Ennylabegan to Enubuj and Kwajalein (FIG. 2 & FIG. 3). Flies are absent on islands without native or American
inhabitants even though adjacent islands may have high densities, which lends
some support to the minimal invasion hypothesis. Fly persistence around human
habitation is probably not to seek breeding sites but rather to attain
foodstuffs, especially carbohydrates, meat scraps and sauces which are
perennially available at recreation sites especially. Therefore, control efforts stress
corralling pigs, reconstructing privies and reducing dog populations. All
residents on all islands are urged to refrain from indiscriminate disposal of
wastes from soft drink and beer cans. Importations of natural enemies are
being stressed for those areas where pigs and dogs roam wild. New Approaches to Bush Fly Reduction New approaches to the solution of the bush fly problem in
Kwajalein atoll involve the use of effective poisoned baits for adult flies,
technological changes in garbage disposal and the importation of natural
enemies from the presumed original endemic area of M. sorbens in central and eastern Africa. Poison Baits Sugar bait mixtures that have been used for M domestica
in years previous to 1972 were wholly ineffective for killing adult M. vetustissima due to their almost
complete lack of attractiveness.
Baits tested during November 1972 through April 1973 included a
variety of decomposing foodstuffs, of which rotting eggs and rotting fish
sauces were very highly attractive. In one experiment using a 6-day old
mixture of one part fresh whole eggs to one part water (Legner 1970b, 1971), it is estimated that about
50,000 bush flies were attracted to the mixture and killed by a 0.5 ppm
Dichlorvos, (8) additive. The poisoned mixture was poured in
quantities of 100 mI each on damp sand at 20 sites in the shade and spaced
every 10m along a public beach on Kwajalein (FIG
6 & FIG 7). Baits placed above the height of 1m or against walls in
open pavilions were only weakly attractive. After 48 hours, flies were
reduced to inconspicuous levels all over Kwajalein Island (Table 1 gives the
approximate % of each species trapped).
This condition endured for 3 consecutive days after which immigrating
flies managed to build up to annoying levels starting on the 4th day when the
baits ceased to be attractive. The former density of flies had not yet been
reached, one week after the baiting experiment; and this population was
subsequently reduced in the same manner by applying additional fresh poisoned
baits. Flies breeding in dog feces were the principal source of the
population recovery on Kwajalein, as shown by marking and recovery
experiments in November 1973. aiting was continued through November 1973 and extended to
other islands in the atoll with a result of sustained reduction of bush fly
to below general annoyance levels (less than 0.01 attracted per minute on
Kwajalein, Roi-Namur, Illeginni and Meck Islands). A new attractant that augmented the rotting egg mixture
consisted of beach sand soaked for one week in the decomposing body fluids of
buried sharks. This new attractant was far superior to rotting eggs both in
rate and time of attraction, the latter sometimes exceeding 5 days. The baiting method of control can be used effectively if
applied initially twice a week. A schedule of biweekly applications or longer
may follow as determined by personnel in charge of the control program. Resistance Resistance in the tropical environment could be expected
to develop very quickly if adult flies that are exposed to the baits are able
to reach breeding sites. This resistance might ultimately be expected to
spread through the whole M. vetustissima
population in Kwajalein atoll, similar to that observed for other species
(Georghiou 1966). Obviously, if breeding sites and poisoned baits are located
together within the normal flight range of flies, resistance rate is
increased. However, there is probably a lower resistance possibility to
electric shock devices, and not much chance for resistance against natural
bait such as rotting eggs or fish.
Nevertheless, with full awareness of the plasticity of M vetustissima and the general
resilience of nature, it cannot be ignored that the possibility for
resistance to any chemical control implemented by humans does exist. Technological
and Cultural Improvements To further ensure
against breeding of M. vetustissima
around refuse disposal sites and that resistance does not develop in
populations of the common house fly and blowfly which are able to
successfully breed in these sites, the development of suitable concrete
barriers to larval flies around such sites was proposed. Fly larvae of M.
domestica and Phaenicia cuprina especially, were repeatedly
observed entering the soil from cans and dumpsters containing refuse. Such
refuse sites placed on adequately rimmed concrete platforms would probably
prevent larvae from finding suitable pupation sites in surrounding soil. The
use of petroleum oils in a long perimeter around refuse deposition sites,
saturating the soil within the depicted circle to a depth of at least 5 cm,
might offer partial control. However, some fly larvae are capable of
penetrating this barrier so that its effectiveness is not as great as
concrete. Natural Enemies Biological control organisms are usually effective only in
one or a few breeding habitats, and their activities are limited under
certain types of climatic stress. For example, when house flies breed in
garbage they are less prone, to attack by certain natural enemies than when
breeding occurs in accumulated animal excrement (Legner and Poorbaugh 1972). The aim of biological
control is to locate and establish natural enemies that will perform
effectively in: all problem breeding habitats. This goal is rarely achieved,
so that effective control takes on an integrated aspect where cultural and
chemical means are implemented. There are no known natural enemies that will specifically
attack the bush fly nor is there any information on what kinds of parasites
and predators range in the droppings of humans, dogs and pigs in eastern
Africa, where bush fly may have originated. The best that can be done in
biological control at this time is to import natural enemies of other fly
species for their possible adaptation to M. vetustissima. (Legner 1970a; Legner and Greathead 1969 ; Legner and Olton 1968, 1970, 1971 ; Legner et al. 1967). This is the approach that has been made in Hawaii and is
partially successful. As new species
of natural enemies become available, they will be introduced for biological
control. There is some expectation that a concerted effort will be made to
seek out specific natural enemies in eastern Africa as support for the work
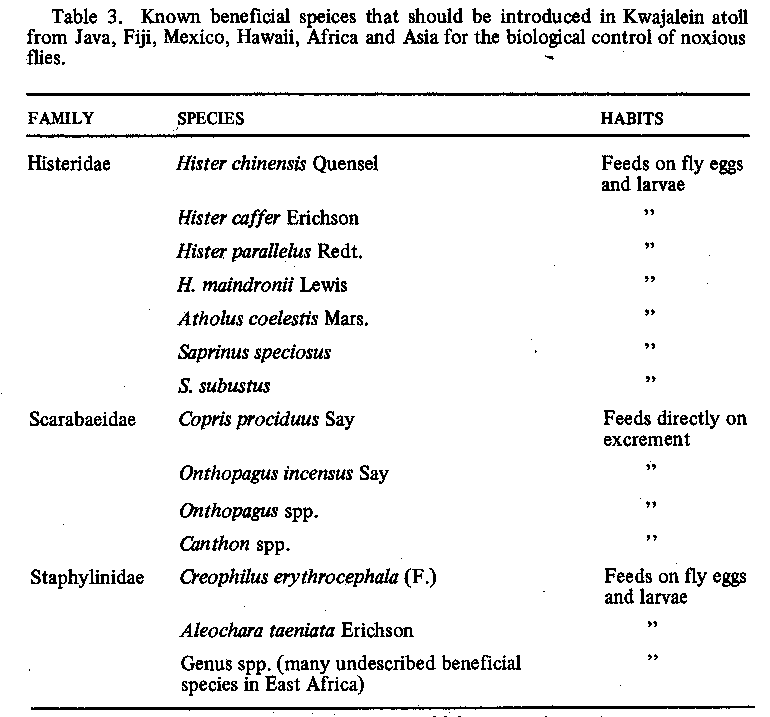
becomes available. A list of species that should be considered for introduction in Kwajalein atoll is
shown in Table 3; however, this list
is expected to grow as information about new natural enemies from Africa
develops. The biological control phase of fly reduction must be
viewed as a long range approach with ever increasing fly reduction as the new
beneficial species become established. An investment of research time in the
initial evaluation of a natural enemy species during the importation phase
would not be practical from the standpoint that such time would, (I) detract from
the amount of effort placed on introductions; (2) most species will probably
not become established judging from a history of biological control efforts
during the past 50 years; and (3) fly densities are expected to drop
gradually with the relatively small numbers of natural enemies being released
at any given time. Once an obvious
drop in the adult M. vetustissima
density has occurred, a thorough evaluation of the natural enemy species
responsible may be carried out. POTENTIAL INVASION OF MUSCA
VETUSTISSIMA IN NORTH AME RICA There has been some speculation about the possibility of Musca
vetustissima invading
portions of North America, where climatic similarities with Africa and
Australia exist. Undoubtedly the more tropical portions of the continent
could sustain this species, with the Florida peninsula and Mexico appearing
the most vulnerable. There is greater uncertainty about its establishment in
the Southwestern United States where winter cold may be an effective barrier.
In Australia M . vetustissima
over-winters with difficulty in the colder southern quarter of the continent
(Norris 1966) and probably reinvades that region annually from the more
tropical north (Hughes 1966). Similarly, in the Middle East Israel seems to
be out of the general range of M. vetustissima
(Legner and Olton 1968), although it is a prominent
species further south along the Nile River (Hafez and Attia 1958). There
being no sustained breeding sites available in the intervening desert,
invasion during warm seasons is precluded. Much the same conditions exist in
the American Southwest where annual invasions from Mexico, a potential
breeding source, may be minimal across the arid Sonoran Desert. The threat of
invasion in the southeastern United States from potential breeding sites in
Florida is greater, however. CONCLUSIONS In modern pest control the importance of being conscious
of what effects a control action may have on the total environment as well as
its permanency are recognized. Resistance to pesticides, especially in the
tropics, is a greater threat in our procedure than pesticide pollution
because we employ compounds that degrade and are nonpoisonous to us. In fly
control on Kwajalein, resistance to Dichlorvos and related compounds would
leave no alternative stop-gap measure to use in case of an epidemic of human
disease in which flies were vectors.
Resistance could also produce hardier strains and theoretically even
more aggressive flies (G. P. Georghiou, personal communication; Legner 1970a). Therefore, it is essential
that the baiting program to combat flies is minimized and that stress be
placed on the implementation of technological, cultural and biological
controls that could lead to a permanent reduction of fly breeding in the
atoll. As of January 2002, the baiting procedure in Kwajalein has
not continued with the sophistication initially determined necessary. The result is that flies are not adequately
reduced where breeding habitat, such as dog feces remain. Personnel changes that resulted in
critical information not being passed on are primarily responsible. Of paramount importance is the proper
brewing of the baits and the latter’s placement in wind-calm areas of the
islands. Also, the baiting program
should logically be extended, at least experimentally, to Musca
vetustissima Walker in Australia, where towns simulate islands in the
vast bush land. Temporary relief from
bush fly annoyance might be expected to occur during summer when these flies
descend into the higher latitudes. ACKNOWLEDGMENTS The assistance and cooperation of personnel in the
following organizations who made this scientific study possible is gratefully
acknowledged: Global Associates,
Department of the Army, University of California, United States Trust
Territory of the Pacific, and the people of the Marshall Islands. REFERENCES CITED [Please
see MELVYL Library for additional references] Awati, P. R., 1920. Bionomics of house-flies. I. Outdoor feeding
habits of house flies with species reference to Musca promiscua (
angustrifrons ? ). J . Med. Res. 7: 548-52. Bell, T. D., 1969. Epidemic glomerulonephritis in Hawaii. Rep.
Pediat. Serv., Dep. Med., Tripler Army Hospital, Honolulu, Hawaii. Mimeo. 25
p. Bevan, L. E. W ., 1926. Report of the Director of Veterinary
Research for the year 1925. Rev. Appl. Ent. (b): 14: 117. Bohart, G. E. and J. L. Gressitt, 1951. Filth-inhabiting flies of
Guam. Bull. B. P. Bishop Museum, Honolulu No.204: 152 p, 17 plates. Bornemissza, G. F., 1970. Insectary studies on the control of
dung breeding flies by the activity of the dung beetle, Onthophagus gazella F
. (Coleoptera: Scarbaeinae). J. Aust. Ent. Soc. 9: 31-41. Bryan, E. H. Jr., 1972. Life in the Marshall Islands. Pacific Scien. Info Center, B. P.
Bishop Museum, Honolulu, Hawaii. 237 p. Buxton, P. A. and G. H. E. Hopkins, 1927. Researches in Polynesia
and Melanesia, Part III, Medical Entomology .Mem. London Sch. Hyg. Trop. Med.
1: 51-85. Gaud, J. and P. Faure, 1951. Effect de la lutte antimouches sur I'insidence des
maladies ocularies dans le sudmarocain. Bull. Soc.
Path. Exot. 44: 446-48. Gaud, J., J. Laurent, and P. Faure, 1954. Biologie de Musca sorbens et role
vecteur probable de cette espece en pathologie humaine au Maroc. Bull. Soc.
Path. Exot. 47: 97-101. Georghiou, G. P ., 1966. Distribution of insecticide-resistant
houseflies on neighboring farms. J. Econ. Ent. 59(2): 341-46. Greenberg, B., 1971. Flies and Disease. Vol. I. Ecology ,
Classification and Biotic Associations. Princeton Univ. Press. Princeton, N
.J .856 p. Hafez, M. and M. A. Attia, 1958. Studies on the ecology of Musca
sorbens Wied. in Egypt. Bull. Soc. Ent. Egypt 42: 83-121. Harris, A. H. and H. A. Down, 1946. Studies of the dissemination
of cysts and ova of human intestinal parasites by flies in various localities
on Guam. Amer. J. Trop. Med. 26: 789-800. Hardy, D. E., 1952. Additions and corrections to Bryan's check
list of the Hawaiian Diptera. Proc. Hawaiian Ent. Soc. 14(3): 443-84. Hughes, R. D., 1968. Bush fly natural history .Rep. Bushfly Res. Sec.,
CSIRO Div. Ent., Canberra, A. C. T . 12 p. Hughes, R. D., 1970. The seasonal distribution of bush fly (Musca
vetustissima Walker) in south-east Australia. J. Anim. Ecol. 39: 691-706. Hughes, R. D. and J. Walker, 1970. The role of food in the population
dynamics of the Australian bush fly. Rep. Div. Ent., CSIRO, Canberra,
Australia. Jack, R. W., 1931. Report of the chief entomologist for the year
1930. Rep. Sec. Dep. Agric. S. Rhodesia, 1930. Rev. Appl. Ent. (B) 19:
128-29. Joyce, C. R., 1950. Notes and exhibitions. Proc. Hawaiian Ent.
Soc. 16(3): 338. Lamborn, W. A., 1937. The haematophagous fly Musca sorbens
Wied., in relations to the transmission of Leprosy. J. Trop. Med. Hyg., 15:
37-42. 1970a Legner, E. F ., 1970a. Comtemporary considerations on the
biological suppression of noxious brachycerous Diptera that breed in
accumulated animal wastes. Proc. Calif. Mosq. Contr. Assoc. 38: 88-89. 1970b Legner, E. F ., 1970b. Attraction of Hippelates eye gnats and
other minute Diptera to baits and man with considerations on competitive
displacement by exotic non-problem species. Proc. Calif. Mosq. Cont. Assoc.
37: 119-26. 1971 Legner, E. F., 1971. Observations on the distribution, relative abundance and behavior
of anthropophilic Chloropidae in the Caribbean area. Carib. J. Sci. 11(3-4):
163-169. 1978 Legner, E. F., 1978. Diptera. Medical and Veterinary Pests. 1012-19; 1043-69. In: C.
P. Clausen [ed.] , "Introduced Parasites and Predators of Arthropod
Pests and Weeds: a Review." U.S. Dept. Agr. Tech. Rept. 1969 Legner, E. F. and D. J.
Greathead, 1969. Parasitism of pupae in East African populations of Musca
domestica and Stomoxys calcitrans. Ann. Ent. Soc. Amer.
62(1): 128-133. 1968 Legner, E. F. and G. S. Olton, 1968. Activity of parasites from
Diptera: Musca domestica, Stomoxys calcitrans, and species of Fannia,
Muscina, and Ophyra II. at sites in the Eastern Hemisphere and Pacific area. Ann. Ent. Soc. Amer. 61(5): 1306-14. 1970 Legner, E. F. and G. S. Olton, 1970. World\vide survey and
comparison of adult predator and scavenger insect populations associated with
domestic animal manure where livestock is artificially congregated. Hilgardia
40(9): 225-66. 1971 Legner, E. F. and G. S. Olton, 1971, Distribution and relative abundance of dipterous pupae and
their parasitoids in accumulations of domestic animal manure in the
southwestern United .States. Hilgardia 40(14): 505-35. 1972
Legner, E. F. and J. H. Poorbaugh, 1972. Biological control of vector and noxious
synanthropic flies: a review. Calif.
St. Dept. Publ. Hlth., Vector Views 19(11): 81-100 1967
Legner, E. F., E. C. Bay, and E. B. White, 1967.
Activity of parasites from Diptera: Musca domestica, Stomoxys calcitrans,
Fannia canicularis and F. femoralis, at sites in the Western
Hemisphere. Ann.
Ent. Soc. Amer. 60(2): 462-68. 1974 Legner, E. F., B. B.
Sugerman, Hyo-sok Yu & H. Lum.
1974. Biological and
integrated control of the bush fly, Musca sorbens Wiedemann and other
filth-breeding Diptera in Kwajalein Atoll, Marshall Islands. Bull Soc. Vector Ecologists (1): 1-14. Lewis, D. J., 1954. Muscidae of medical interest in the
Anglo-Egyptian Sudan. Bull. Ent. Res. 45: 783-96. McGuire, C. D. and R. C. Durant, 1957. The role of flies in the transmission of eye disease in Egypt.
Amer. I. Trop. Med. Hyg. 6: 569-75. Meng, C. and G. F. Winfield, 1938. Studies on the control of fecal-borne diseases of North China.
V. A preliminary study of the density , species make-up, and breeding habits
of house frequenting fly populations of Tsinan, Shantung, China. Chinese Med.
I. Suppl. II: 463-86. Meng, C. H. and G. F. Winfield, 1944. Breeding habits of common West China flies. Chinese Med. I.
62(A): 77-87. Norris, K. R., 1966. Notes on the ecology of bush fly, Musca
vetustissima Walk. (Diptera: Muscidae) in the Canberra District. Aust. I. Zool. 14: 1139-56. Patterson, H. E. and K. R. Norris, 1970. The Musca sorbens
complex: the relative status of the Australian and two African populations. Aust. I. Zool. 18: 231-45. Patton, W. S., 1922. Notes on the species of the genus Musca,
Linnaeus -Part I. Bull. Ent. Res. 12: 411-26. Patton, W. S., 1923. Some Philippine species of the genus Musca,
Linnaeus. Philipp. I. Sci. 23: 309-22. Patton, W. S., 1926. The Ethiopian species of the genus Musca,
L. Rec.Indian Mus. 28: 29-52. Patton, W. S., 1932. Studies on the higher Diptera of medical and
veterinary importance: a revision of the species of genus Musca, based on a
comparative study of the male terminalia. I. The natural grouping of the
species and their relationship to each other. Ann. Trop. Med. parasit. 26:
347. Patton, W. S.,1933. Studies on the higher Diptera of medical and
veterinary importance: a revision of the species of the genus Musca, based on
a comparative study of the male terminalia. II: A practical guide to the Palearctic
species. Ann. Trop. Med. Parasit. 27: 327-45, 397-430. Pat ton, W. S. 1936. A revision of the species of genus Musca
based OIl a comparative study of the male terminalia. III. A practical guide to the Ethiopian
species. Ann. Trop. Med. Parasit. 30: 469-90. Peffly, R. L., 1953a. A summary of recent studies on houseflies
in Egypt. J. Egypt. Pub1. I-llth. Assn. 28: 55-74. Peffly, R. L. 1953b. The relative importance of different fly
breeding materials in an Egyptian village. J. Egypt. Publ. I-llth. Assn. 28:
167-80. Roy, D. N. and L. B. Siddons, 1940. On continuous breeding of
flies in the laboratory .Ind. J. Med. Res. 28: 621-24. Sabrosky, C. W ., 1952. Houseflies in Egypt. Amer. J. Trop. Med, Hyg. 1 : 333-36. Smirov, E. S., 1940. Le probleme des mouches a Tadjikistane. Medskaya
Parasit. 9: 515-17. Thomson, J. C. and W. A. Lambom, 1934. Mechanical transmission of
trypanosomiasis and yaws through the agency of non-biting haematophagous
flies. (Preliminary note on experiments). Brit. Med. J. 2845: 506-09. Van Emden, F. I., 1965. The fauna of India and the adjacent countries. Diptera Vol. 7., Muscidae,
Pt. I. Gov. Publ. India, Delhi, India. Wiedemann, C. R. W ., 1830.
Aussereurop. Zweifl. Insekt.lI: 1-684. Wilton, D. P ., 1963. Dog excrement as a factor in community fly
problems. Proc. Hawaiian Ent. Soc. 28(2): 311-17. Yu, Hyo-sok, 1971. The biology and public health significance of Musca sorbens
Wied. in Hawaii. M. s. Thesis, Univ.
of Hawaii. 72 p. ---------
---------- (1) This study was supported in part by grants- in-aid and
assistance of Global Associates, Oakland, California. (2) Professor of Biological
Control Emeritus, University of California, Riverside 92521. (3)
Entomologist, Trust Territory of The Pacific (4) Research Assistant, University of
California, Riverside 92502. (5) Foreman, Facilities,
Global Associates, Kwajalein, APO San Francisco 96555 (6) Ratio of the least
width of the frons and the greatest width of the head. (7) Estimates of Musca vetustissima
adult density were made between 10-1500 hours by
counting the number of alightments per minute on the face, neck and ear areas
of the body while standing upright in the shade on the rainless days. At
least 20 such estimates were gathered on anyone island per day and the
results averaged for text presentation. sources, with natural enemies playing
a secondary role. (8) 2,2-dichlorovinyl
dimethly phosphate |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}